I
ESTRUTURAS VESTIGIAIS
Gerd B. Müller
University of Vienna
Oxford University Press, 2002
Órgãos e estruturas vestigiais (também chamados de vestigia, rudimentos ou remanescentes) são partes ou órgãos do corpo reduzidos, muitas vezes sem função visível nas espécies portadoras derivadas, que foram totalmente desenvolvidos e funcionais em membros anteriores (ancestrais) dessa linhagem filogenética. Essas estruturas, às vezes descritas como atrofiadas ou degeneradas, são geralmente pequenas em comparação com seu tamanho relativo em gerações ancestrais ou em espécies próximas.
Vestigia, é uma palavra etimologicamente derivada do latim vestigium (pegada, traço, marca), e desempenhou um papel importante na fundação da teoria evolutiva porque representava traços tangíveis de gerações passadas em organismos recentes. Hoje, os vestígios anatômicos não representam um tópico importante na pesquisa evolutiva, sua presença em todas as espécies é tida como certa, embora os mecanismos causais responsáveis tanto por sua redução quanto por sua retenção não sejam totalmente compreendidos.
As estruturas vestigiais são identificadas pelo método comparativo. Isso significa que a homologia (semelhança, identidade) do caráter reduzido com uma contraparte ancestral mais desenvolvida deve ser estabelecida não apenas com base em critérios estruturais e posicionais, mas também com relação à presença contínua da estrutura na linhagem que conduz do ancestral para a forma derivada. [Os órgão homólogos são aqueles que podem ou não realizar a mesma função, porém apresentam uma estrutura básica igual, mesma origem e desenvolvimento embrionário. Isso quer dizer, portanto, que os indivíduos que apresentam esses órgãos possuem ancestralidade compartilhada.] O último critério representa uma distinção crítica de características chamadas atavismos, que ocorrem após períodos prolongados de ausência completa de uma característica.
Enquanto os atavismos aparecem apenas em casos excepcionais e em indivíduos isolados, uma estrutura vestigial está presente em todos os representantes de uma espécie. Outra distinção deve ser feita com relação ao termo rudimento. Embora muitas vezes usado como sinônimo de vestigium, um rudimento é propriamente um primórdio de desenvolvimento ou a base rudimentar de um órgão em particular, especialmente no embrião, e coincide com um vestigium apenas quando um primórdio é retido no adulto.
A vestigialização afeta estruturas e órgãos que reduziram ou perderam sua função como resultado de uma mudança evolutiva no estilo de vida das formas derivadas. Exemplos clássicos são as reduções de olhos e membros em organismos que vivem em cavernas ou que se enterram, reduções de asas associadas à habitação no solo ou reduções dentárias e intestinais atribuíveis a mudanças na dieta.
Os órgãos afetados podem apresentar diferentes estágios de redução em diferentes membros de um clado. Assim, vemos diferentes graus de redução de membros em lagartos, ossos pélvicos de tamanhos diferentes em certas baleias e serpentes, e vários estágios de regressão ocular em organismos cavernosos vertebrados e não vertebrados, ou de rudimentos de asas em pássaros e insetos que não voam.
Em muitos casos, os vestígios estão presentes apenas em formas fósseis e são completamente perdidos em representantes existentes de uma linhagem. Em outros casos, os rudimentos estão presentes apenas no embrião ou na fase juvenil e são totalmente perdidos no adulto, como os dentes nos embriões das baleias de barbatana. [poderíamos dizer que a presença de notocorda no embrião de cortados e sua retenção em humanos até quatro anos é um traço vestigial. A notocorda é formada por sinais indutores, vindos da região da linha primitiva, que induzem as células precursoras notocordais a formá-la. A notocorda tem como funções definir o eixo do embrião, serve de base para a formação do esqueleto axial, e é o futuro local dos corpos das vértebras.]
Extensas coleções de estruturas vestigiais de muitos organismos e de uma ampla gama de sistemas de órgãos podem ser encontradas em tratados clássicos de morfologia comparativa. Wiedersheim (1893), por exemplo, fornece longas listas de casos na anatomia humana, incluindo o freqüentemente citado apêndice vermiforme, a glândula pineal, o órgão vômero-nasal, o osso coccígeo, os músculos da orelha, os terceiros dentes molares, restos de ductos néfricos e genitais, e muitos mais. O grau em que os órgãos vestigiais são desenvolvidos no adulto pode variar extensivamente dentro e entre as espécies. Frequentemente, as estruturas são relativamente mais desenvolvidas no embrião do que no adulto. Quando sua presença está confinada ao embrião, a ideia de recapitulação é evocada.
Os mecanismos evolutivos da vestigialização são considerados bem compreendidos. Argumenta-se que mudanças comportamentais ou ambientais iniciais levam a uma perda parcial ou total da função de um órgão, com consecutiva seleção contra essa estrutura, o que teria se tornado um fardo ou obstáculo para evolução posterior. No entanto, até que ponto a vestigialização pode ser explicada em termos de adaptação é debatida. A seleção direta para vestigialização parece ser rara. Em vez disso, os órgãos geralmente sofrem redução como subproduto de outras mudanças adaptativas, como a redução de membros em associação com o alongamento ou miniaturização do corpo. Outras explicações invocam a seleção indireta por meio de trocas de energia ou pleiotropia antagônica. Mas os cenários não adaptativos prevalecem: as estruturas podem degenerar com mais frequência devido a um relaxamento da seleção estabilizadora e ao acúmulo de mutações seletivamente neutras.
A realização específica e desenvolvimental da vestigialização pode ser efetuada por vários processos. Alterações mutacionais podem afetar o controle do desenvolvimento, perturbar a divisão celular e a diferenciação histológica ou interromper as interações indutivas. Vários modos gerais podem ser distinguidos. Ou toda a progressão de um primórdio embrionário é retardada ou interrompida, levando à preservação de um rudimento no adulto; ou o desenvolvimento de um órgão prossegue normalmente até certo ponto em que é ativamente destruído, por exemplo, por atividade hormonal, apoptose ou fagocitose, como na cauda de girinos anuros. Uma terceira possibilidade é que mudanças gerais e heterocrônicas no desenvolvimento larval ou embrionário simplesmente impedem que as estruturas adultas se formem completamente. Como regra geral, a vestigialização morfológica ocorre na ordem inversa da sequência do desenvolvimento normal. Por exemplo, dígitos de membros reptilianos em regressão são deletados na ordem inversa de sua formação embrionária, redução ocular de peixes e salamandras que vivem em cavernas segue uma progressão distal para central da córnea, para o cristalino, para o bulbo e para os componentes nervosos, aproximadamente a ordem oposta de sua formação. [eye reduction of cave-dwelling fish and sal. amanders follows a distal to central progression from cornea, to lens, to bulbus, to nervous components-roughly the opposite order of their formation.]
A questão mais interessante da vestigialização provavelmente diz respeito não ao fato da regressão estrutural em si, mas sim à manutenção de remanescentes que não têm função óbvia nos portadores derivados, ou mesmo parecem estar sob seleção negativa. Um exemplo é o apêndice vermiforme, que deve ter causado grande número de mortes pré-reprodutivas durante a evolução dos hominídeos. Como é bastante improvável que estruturas completamente sem função sejam preservadas, certas funções devem ser assumidas como persistentes. Uma classe óbvia de tais funções é seu papel intermediário no desenvolvimento, quando uma estrutura fortemente integrada nas interações de desenvolvimento não pode ser removida sem efeitos deletérios posteriores (a jusante). [The most interesting question of vestigialization probably concerns not the fact of structural regression itself, but rather the maintenance of remnants that have no obvious function in the derived bearers, or even seem to be under negative selection. An example is the vermiform appendix, which must have caused extensive numbers of prereproductive deaths during hominid evo-lution. Since it is quite unlikely that structures com. pletely without function would be preserved, certain functions must be assumed to persist. One obvious class of such functions is their intermediate role in develop-ment, when a structure that is tightly integrated in de velopmental interactions cannot be removed without deleterious downstream effects.]
Em outros casos, as estruturas vestigiais podem ter adquirido funções novas, menos óbvias e diferentes das originais. Portanto, um vestigium geralmente não deve ser considerado sem função, ou apenas com relação a seus papéis ancestrais e adultos.
Finalmente, estruturas vestigiais (vestigia) podem ser mantidas por causa de redes de genes intactas. A redução ou perda de uma estrutura fenotípica não significa necessariamente que sua base genética também esteja perdida. Mesmo os genes estruturais que não foram ativados por muitos milhões de anos, como os genes das proteínas do esmalte em aves ou as proteínas do cristalino em toupeiras, são frequentemente retidos no genoma.
O significado das estruturas vestigiais, esses ecos do passado, reside hoje principalmente na ilustração de origens não adaptativas de certas características da construção do organismo. Eles também são de interesse taxonômico, e uma série de fenômenos morfológicos só podem ser compreendidos se os processos de vestigialização forem conhecidos. É o caso dos atavismos, muitas vezes baseados em vestígios embrionários, como os ocasionais dedos extras em cavalos. Outro caso é a inovação morfológica, que pode ter origem em rudimentos retidos de estruturas reduzidas. Aprimoramentos experimentais de estruturas vestigiais embrionárias também podem revelar interações de desenvolvimento suprimidas. Finalmente, certos vestígios em humanos são de importância médica, como exemplificado por cistos paraovarianos, fístulas branquiais ou apendicite. Em geral, as estruturas vestigiais e os processos de sua formação lançam luz sobre a relação entre desenvolvimento e evolução [especialmente sobre a macroevolução, i.e., sobre a origem de táxons superiores a espécie.]
II
Um órgão vestigial é um órgão que perdeu sua função original ao longo do tempo. Órgãos vestigiais podem ser encontrados em plantas e animais.
Alguns órgãos vestigiais em humanos
Menbraba nictitante (placa semilunaris)
Musculos para mover a orelha
Caninos pontudos
Pelos no corpo
Terceiro molar
Mamas nos machos
Apendice vermiforne
Musculos piramidalis
Vestigio caudal (coxis)
Segmentação dos músculos abdominais
Órgãos vestigiais em plantas
Nas plantas, os órgãos vestigiais são freqüentemente chamados de “folhas vestigiais”. São folhas que perderam sua função original com o tempo e não são mais utilizadas para fotossíntese ou outros fins. Folhas vestigiais podem ser encontradas nos caules de algumas plantas e muitas vezes parecem folhas pequenas e achatadas.
Nossa compreensão dos órgãos vestigiais evoluiu ao longo do tempo à medida que novas informações se tornaram disponíveis.
No século VXIII, acreditava-se que cada órgão do corpo tinha uma função. No entanto, à medida que a ciência progrediu, descobrimos que alguns órgãos não servem mais a um propósito. Órgãos vestigiais são restos de órgãos que perderam sua função ao longo do tempo. Eles podem ser encontrados em plantas e animais.
Nas plantas, os órgãos vestigiais podem assumir muitas formas diferentes. Por exemplo, algumas plantas têm folhas pequenas e rudimentares que não servem mais para nenhuma função. Outras têm pétalas reduzidas em tamanho ou número. Algumas plantas ainda têm raízes vestigiais que não funcionam mais.
Existem várias explicações possíveis para a evolução dos órgãos vestigiais nas plantas. Uma possibilidade é que o órgão já foi necessário para a sobrevivência, mas com o tempo tornou-se menos importante. Isso pode ser devido a mudanças no ambiente ou à evolução de novos órgãos mais eficientes. Outra possibilidade é que o órgão nunca foi necessário em primeiro lugar e é simplesmente uma relíquia da evolução.
Apesar de sua falta de função, os órgãos vestigiais ainda podem desempenhar um papel importante no desenvolvimento da planta. Por exemplo, eles podem ajudar a definir a forma e a estrutura da planta. Eles também podem influenciar a forma como a planta cresce e se reproduz.
No geral, os órgãos vestigiais são um exemplo interessante de como a evolução pode fazer com que os órgãos percam sua função ao longo do tempo. Embora possam não servir mais a um propósito, eles ainda podem desempenhar um papel importante no desenvolvimento da planta.
III
Na obra de Charles Darwin's On the Origin of Species (1859) Sobre a Origem das Espécies, e em seus trabalhos posteriores, ele se referiu a várias estruturas “vestígiais” na anatomia humana que sobraram do curso da evolução. Esses órgãos vestigiais, argumentou Darwin, são evidências da evolução e representam uma função que já foi necessária para a sobrevivência, mas com o tempo essa função tornou-se diminuída ou inexistente.
A presença de um órgão em um organismo que se assemelha a um encontrado em outro levou os biólogos a concluir que esses dois podem ter compartilhado um ancestral comum. Órgãos vestigiais demonstraram de forma notável como as espécies estão relacionadas umas com as outras filogeneticamenge e forneceram um terreno sólido para a ideia de descendência comum se sustentar. Da descendência comum, prevê-se que os organismos retenham esses órgãos vestigiais como remanescentes estruturais de funções perdidas. É apenas por causa da teoria macroevolucionária, ou evolução que ocorre em períodos muito longos de tempo, que esses vestígios aparecem.
VESTIGIAL ORGANS AND STRUCTURES
Gerd B. Müller
University of Vienna
Vestigial organs and structures (also called vestigia, rudiments, or remnants) are reduced body parts or organs, often without visible function in the derived bearers, that were fully developed and functioning in earlier members of that phylogenetic lineage. These structures, sometimes described as atrophied or degenerate, are usually small in comparison with their relative size in ancestral generations or in closely related species.
Vestigia, etymologically derived from Latin vestigium (footprint, trace, mark), played an important role in the founding of evolutionary theory because they represented tangible traces of past generations in recent organisms. Today, anatomical vestigia do not represent a major topic in evolutionary research, their presence in all species is taken as given, although the causal mechanisms responsible for both their reduction and their retention are not fully understood.
Vestigial structures are identified by the comparative method. This means that homology (sameness, identity) of the reduced character with a more fully developed, ancestral counterpart must be established not only on the basis of structural and positional criteria but also with regard to the continuous presence of the structure in the lineage leading from the ancestral to the derived form. The latter criterion represents a critical distinction from features called atavisms, which occur after prolonged periods of complete absence of a character.
Whereas atavisms appear only in exceptional cases and in single individuals, a vestigial structure is present in all representatives of a species. Another distinction should be made with regard to the term rudiment. Although often used synonymously with vestigium, a rudiment is properly a developmental primordium or anlage, and it coincides with a vestigium only when a primordium is retained in the adult. (anlage the rudimentary basis of a particular organ or other part, especially in an embryo.)
Vestigialization affects structures and organs that have reduced or lost their function as the result of an evolutionary change in lifestyle of the derived forms. Classical examples are eye and limb reductions in cave-dwelling or burying organisms, wing reductions associated with ground dwelling, or dental and intestinal reductions attributable to changes of diet. The affected organs can show different stages of reduction in different members of a clade. Thus, we see different degrees of limb reduction in skinks, pelvic bones of different sizes in certain whales and snakes, and various stages of eye regression in vertebrate and nonvertebrate cave organisms, or of wing rudiments in flightless birds and insects. In many cases, the vestigia are present only in fossil forms and are completely lost in extant representatives of a lineage. In other cases, the rudiments are present only in the embryo or juvenile and are completely lost in the adult, such as teeth in the embryos of baleen whales.
Extensive collections of vestigial structures from many organisms and from a wide range of organ systems can be found in classical treatises of comparative morphology. Wiedersheim (1893), for example, provides long lists of cases in human anatomy, including the often cited vermiform appendix, the pineal gland, the vomero-nasal organ, the coccygeal bone, muscles of the ear, the third molar teeth, remnants of nephric and genital ducts, and many more. The degree to which vestigial organs are developed in the adult can vary extensively both within and among species. Often the structures are relatively more developed in the embryo than in the adult.
When their presence is confined to the embryo, the idea of recapitulation is evoked.
The evolutionary mechanisms of vestigialization are thought to be well understood. It is argued that initial behavioral or environmental changes lead to a partial or complete loss of function of an organ, with consecutive selection against that structure, which would have become a burden or hindrance for further evolution. Yet the extent to which vestigialization can be explained in terms of adaptation is debated. Direct selection for vestigialization seems to be rare. Rather, organs often undergo reduction as a byproduct of other adaptive changes, such as limb reduction in association with body elongation or miniaturization. Other explanations invoke indirect selection through energy trade-offs or antagonistic pleiotropy. But nonadaptive scenarios pre-vail: structures may most often degenerate because of a relaxation of stabilizing selection and the accumulation of selectively neutral mutations.
The specific, developmental realization of vestigialization can be effected by various processes. Mutational changes can affect developmental control, disturb cell division and histological differentiation, or interrupt inductive interactions. Several general modes can be distinguished. Either the entire progression of an embryonic primordium is slowed down or halted, leading to the preservation of a rudiment in the adult; or development of an organ proceeds rather normally up to a certain point at which it is actively destroyed for example, by hormonal activity, apoptosis, or phagocytosis, as in the tail of anuran tadpoles. A third possibility is that general, heterochronic shifts in larval or embryonic development simply prevent adult structures from forming fully. As a rule of thumb, morphological vestigialization proceeds in the reverse order of the sequence of normal development. For instance, digits of regressing reptilian limbs are deleted in the reverse order of their embryonic formation; eye reduction of cave-dwelling fish and salamanders follows a distal to central progression from cornea, to lens, to bulbus, to nervous components-roughly the opposite order of their formation.
The most interesting question of vestigialization probably concerns not the fact of structural regression itself, but rather the maintenance of remnants that have no obvious function in the derived bearers, or even seem to be under negative selection. An example is the vermiform appendix, which must have caused extensive numbers of prereproductive deaths during hominid evolution. Since it is quite unlikely that structures completely without function would be preserved, certain functions must be assumed to persist. One obvious class of such functions is their intermediate role in development, when a structure that is tightly integrated in developmental interactions cannot be removed without deleterious downstream effects. In other cases, vestigial structures may have acquired new, less obvious functions that differ from the original ones. Hence, a vestigium should not generally be considered without function, or only with respect to its ancestral, adult roles.
Finally, vestigia may be maintained because of intact gene networks. Reduction or loss of a phenotypic structure does not necessarily mean that its genetic basis is lost too. Even structural genes that have not been activated for many millions of years, such as those for enamel proteins in birds or lens proteins in moles, are often retained in the genome.
The significance of vestigial structures, these echoes of the past, today lies mostly in the illustration of non-adaptive origins of certain features of organismal construction. They are also of taxonomic interest, and a number of morphological phenomena can be understood only if the processes of vestigialization are known.
This is the case with atavisms, which are often based on embryonic vestigia, such as the occasional extra toes in horses. Another case is morphological innovation, which can take its origin from retained rudiments of reduced structures. Experimental enhancements of embryonic vestigia can also uncover suppressed develop mental interactions. Finally, certain vestigia in humans are of medical significance, as exemplified by paraovarian cysts, branchial fistulae, or appendicitis. In general, vestigial structures and the processes of their formation shed light on the relationship between development and evolution.
IV
A existência de estruturas vestigiais é uma das principais linhas de evidência da macroevolução. Aqui eu apresento uma abordagem de agrupamento filogenético para a identificação de estruturas vestigiais e a aplico a Dinosauria. De acordo com essa abordagem, uma estrutura é considerada vestigial se, em comparação com seu homólogo em pelo menos três outgroups sucessivos, for reduzida a um terço ou menos de seu tamanho em relação às estruturas adjacentes e se pelo menos distalmente tiver perdido a morfologia especializada presentes nos três grupos externos.
Esta abordagem identifica:
1) Os dedos IV e V como vestigiais em dinossauros em geral, II-V em saurópodes, III em Tyrannosauridae e Caudipteryx, II e III em Shuvuuia e I e III em pássaros modernos.
2) Todo o membro anterior distal ao cotovelo é vestigial em Abelisauridae.
3) As partes vestigiais da cintura pélvica e dos membros posteriores incluem a haste púbica em Iguanodontia e Ceratopsia,
4) Todo o púbis em Ankylosauria,
5) O primeiro metatarso em Iguanodontia derivados,
6) O primeiro eixo metatarsal em Theropoda e
7) O quinto dedo em dinossauros em geral.
8) Os Centrosaurinae derivados e alguns Chasmosaurus exibem chifres supraorbitais vestigiais.
9) Alguns centrosaurines têm um chifre nasal vestigial.
10) A cavidade antorbital é vestigial em Thyreophora, Iguanodontia e Ceratopsidae. Recomendo que esta informação seja explorada para aumentar a conscientização pública sobre as evidências da macroevolução.
(The existence of vestigial structures is one of the main lines of evidence for macroevolution. Here I introduce a phylogenetic bracketing approach to the identification of vestigial structures and apply it to Dinosauria. According to this approach, a structure is considered vestigial if, in comparison with its homolog in at least three successive outgroups, it is reduced to one-third or less its size relative to adjacent structures and if at least distally it has lost the specialized morphology present in the three outgroups. This approach identifies fingers IV and V as vestigial in dinosaurs in general, II–V in sauropods, III in Tyrannosauridae and Caudipteryx, II and III in Shuvuuia and I and III in modern birds. The entire forelimb distal to the elbow is vestigial in Abelisauridae. Vestigial parts of the pelvic girdle and hindlimb include the pubic shaft in Iguanodontia and Ceratopsia, the entire pubis in Ankylosauria, the first metatarsal in derived Iguanodontia, the first metatarsal shaft in Theropoda and the fifth toe in dinosaurs in general. Derived Centrosaurinae and some Chasmosaurus exhibit vestigial supraorbital horns. Some centrosaurines have a vestigial nasal horn. The antorbital cavity is vestigial in Thyreophora, Iguanodontia and Ceratopsidae. I recommend that this information be exploited to increase public awareness of the evidence for macroevolution. (zslpublications))
Embrião na Terceira Semana
O embrião na caracteriza-se por:
Aparecimento da linha primitiva
Formação da notocorda
Formação do disco trilaminar
1) Gastrulação: Formação das camadas germinativas
Processo pelo qual o disco embrionário bilaminar é convertido em disco embrionário trilaminar (inicio da morfogênese). Durante a gastrulação ocorrem alguns eventos importantes como a formação da linha primitiva, camadas germinativas, placa precordal e notocordal. Cada uma das três camadas germinativas dará origem a tecidos e órgãos específicos:
Ectoderma: Origina a epiderme, sistema nervoso central e periférico e a várias outras estruturas;
Mesoderma: Origina as camadas musculares lisas, tecidos conjuntivos, e é fonte de células do sangue e da medula óssea, esqueleto, músculos estriados e dos órgãos reprodutores e excretor;
Endoderma: Origina os revestimentos epiteliais das passagens respiratórias e trato gastrointestinal, incluindo glândulas associadas.
Formação da Linha Primitiva
No início da terceira semana a linha primitiva surge na extremidade caudal do embrião como resultado da proliferação e migração de células do epiblasto para o plano mediando do disco embrionário, constituindo o primeiro sinal da gastrulação. Na sua extremidade cefálica surge o nó primitivo, com uma pequena depressão no centro chamado fosseta primitiva e ao longo da linha forma-se o sulco primitivo. O aparecimento da linha primitiva torna possível identificar o eixo embrionário.
Após esse processo, ocorre a invaginação de células do epiblasto que dão origem as três camadas germinativas do embrião: o mesênquima ou mesoblasto, que origina os tecidos de sustentação e conjuntivos do corpo, um pouco forma o mesoderma intra-embrionário e outras deslocam o hipoblasto e formam endoderma intra-embrionáiro. As demais células que permanecem no epiblasto formam o ectoderma intra-embrionario. A linha primitiva regride e desaparece na quarta semana do desenvolvimento.
Formação do processo notocordal
Células mesenquimais migram cefalicamente do nó e da fosseta primitiva formando um cordão celular mediano o processo notocordal. Esse processo adquire uma luz - canal notocordal, e cresce até alcançar a placa precordal, área de células endodérmicas firmemente aderidas a ectoderma. Estas camadas fundidas formam a membrana bucofaríngea (boca). Caudalmente a linha primitiva há uma área circular também com disco bilaminar, a membrana cloacal (ânus).
A notocorda surge pela transformação do bastão celular do processo notocordal. O assoalho do processo notocordal funde-se com o endoderma e degeneram. Ocorre então a proliferação de células notocordais a partir da extremidade cefálica, a placa notocordal se dobra e forma a notocorda.
A notocorda:
Define o eixo do embrião;
Base para formação do esqueleto axial;
Futuro local dos corpos vertebrais.
Formação do Alantóide
O alantóide é um anexo embrionário que surge por volta do 16° dia na parede caudal do saco vitelino. Durante a maior parte do desenvolvimento, o alantóide persiste como uma linha que se estende da bexiga urinária até a região umbilical, chamada de úraco, a qual nos adultos corresponderá ao ligamento umbilical mediano.
2) Neurulação: Formação do tubo neural
A formação da placa neural é induzida pela notocorda em desenvolvimento. Por volta do 18° dia, a placa neural se invagina ao longo do eixo central, formando o sulco neural mediano, com pregas neurais em cada lado. No fim da terceira semana, as pregas neurais começam a aproximar-se e a se fundir, formando o tubo neural, primórdio do SNC. Este logo se separa do ectoderma da superfície, se diferencia e forma a epiderme da pele. A fusão das pregas neurais avança em direção cefálica e caudal, permanecendo abertas na extremidade cranial, neuroporo rostral, até o 25º dia e na extremidade caudal, neuroporo caudal, até o 27º dia. Concomitante a esse processo, as células da crista neural migram e formam uma massa entre o ectoderma e o tubo neural, a crista neural. Logo, a crista se separa em duas partes, direita e esquerda, e origina os gânglios espinhais e os gânglios do sistema autônomo e as meninges.
3) Desenvolvimento dos somitos
Durante a formação da notocorda e do tubo neural, o mesoderma intra-embrionário se divide em: mesoderma paraxial, intermediário e lateral (contínuo com o mesoderma extra-embrionário). Próximo ao fim da 3° semana de gestação, o mesoderma paraxial diferencia-se e forma os somitos. No fim da 5° semana 42 a 44 pares de somitos estão presentes e avançam cefalocaudalmente dando origem à maior parte do esqueleto axial e músculos associados, assim como a derme da pele adjacente.

Desenvolvimento do tubo neural por meio do fechamento e fusão das pregas neurais (setas). Com destaque para a diferenciação do mesoderma intra-embrionário. Em a) e b) embrião de 20 dias de desenvolvimento, em c) e d) embrião de 22 dias de desenvolvimento (Rohen & Lutien-Drecoll (2005).

Esquema ilustrativo da formação do tubo neural por meio da fusão das pregas neurais (Moore and Persaud, 2003).
4) Desenvolvimento do celoma intra-embrionário
No interior do mesoderma lateral e cardiogênico surgem espaços celômicos que se unem e formam o celoma intra-embrionário, dividindo o mesoderma lateral em duas camadas:
Camada parietal/ somática que cobre o âmnio;
Camada visceral/ esplâncnica que cobre o saco vitelino:
Somatopleura = mesoderma somático + ectoderma sobrejacente
Esplancnopleura = mesoderma esplacnico + endoderma subjacente
Durante o 2° mês, o celoma está dividido em 3 cavidades:
Cavidade pericárdica;
Cavidades pleurais;
Cavidade peritoneal.

Desenvolvimento dos sólitos e celoma intra-embrionário
(Moore and Persaud, 2003)
5) Desenvolvimento do sistema cardiovascular
No inicio da 3°semana começa a angiogênese no mesoderma extra-embrionário do saco vitelino, do pedículo do embrião e do córion. A formação dos vasos sanguíneos inicia-se com a agregação dos angioblastos, ilhotas sanguíneas. Pequenas cavidades vão se formando dentro das ilhotas, os angioblatos se achatam e originam o endotélio primitivo. Essas cavidades se unem formando redes de canais endoteliais.
O coração e os grandes vasos provêm de células mesenquimais da área cardiogênica. Durante a 3° semana os tubos endocárdicos se fundem, originando o tubo cardíaco primitivo. No fim da 3° semana o sangue já circula e desenvolve-se o primórdio de uma circulação uteroplacentária.
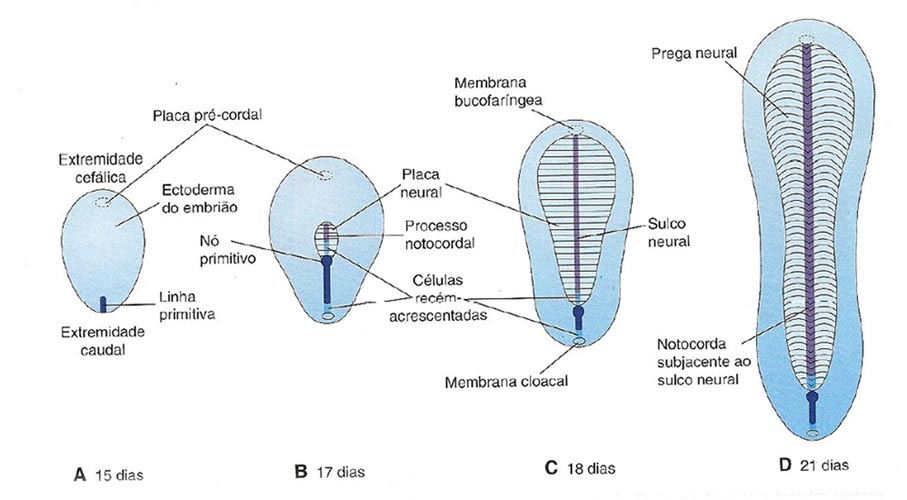
Vistas dorsais do disco embrionário, mostrando como ele se alonga e muda de forma durante a terceira semana (Moore and Persaud, 2008)
Fontes
Para uma visão alternativa
Bibliografia embriologia
Moore KL, Persaud TVN. Embriologia clínica. 8a ed. Rio de Janeiro (RJ): Elsevier; 2008.
Rohen JW, Lutjen-Drecoll E. Embriologia funcional: o desenvolvimento dos sistemas funcionais do organismo humano. 2a ed. Rio de Janeiro (RJ): Guanabara Koogan; 2005.
O’Rahilly R, Müller F. Embriologia & teratologia humanas. 3a ed. Rio de Janeiro (RJ): Guanabara Koogan; 2005.
Schoenwolf GC, Bleyl SB, Brauer PR, Francis-West PH. Larsen’s human embryology. 4th ed. Philadelphia: Churchill & Livingston; 2008.
Moore KL, Persaud TVN. The developing human: clinically oriented embryology. 7th ed. Philadelphia: WB Saunders; 2003.
Sadler TW. Langman embriologia médica. 9a ed. Rio de Janeiro (RJ): Guanabara Koogan; 2005.
Nenhum comentário:
Postar um comentário