Filo CHORDATA
Subfilo UROCHORDATA
(TUNICATA)


Urocordado adulto

Larva de urocordado ou tunicado

Urocordado: estrutura corporal do adulto e da larva

Clavelina moluccensis um urocordados = tunicado
da Australia, pacifico ocidental,Papua, New Guinea...)
(clavelina pequeno bujão ou garrafinha)
(By Nhobgood)



Músculos larvais da cauda e comportamento natatório.
a) Diagrama de um larva Ciona robusta, mostrando o arranjo de 18 células musculares mononucleadas em uma faixa muscular em um lado da cauda.
b) Imagens sobrepostas tiradas em intervalos de 5 milissegundos, mostrando metade de uma batida de cauda no repetitivo comportamento natatório do "girino" Ciona.
c) Ilustração de uma célula muscular da cauda na larva do Aplidium constellatum, mostrando a posição oblíqua das miofibrilas em relação ao eixo anterior – posterior da célula, e a natureza contínua das fibras estriadas de célula para célula.
d) Imagem de Diplosoma listerianum cauda de larva corada com faloidina-Alexa Fluor 488 (miofbrilas, roxo) e DAPI (núcleos, laranja). a e b Adaptado de Nishino et al. [244]. c Adaptado de Grave.


Estrutura de uma Ascída típica

Colônia de tunicados
Os tunicados, ou urocordados, são um subfilo do filo Chordata, que compartilham com outros membros do filo as seguintes características:
(1) uma notocorda permanente ou temporária, na forma de uma haste ou bastão dorsal;
(2) um sistema nervoso central, na forma de um tubo nervoso oco dorsal;
(3) uma faringe provida de fendas branquiais ou bolsas faríngeas e uma glândula ventral secretando iodoproteínas (endóstilo ou tireóide); e
(4) uma cauda muscular pós-anal.
Os tunicados são considerados o grupo irmão dos vertebrados, formando com os últimos o clado Olfactoria. Recentemente, foi proposto classificá-los como um filo dentro do superfilo Chordata. Eles são tradicionalmente subdivididos em três classes:
1) Ascidiacea (bentônica e séssil),
2) Thaliacea, e
3) Larvacea ou Appendicularia (pelágica).
Ascidiacea
São os cordados invertebrados mais conhecidos dentro dos subfilo Urochordata. Como todo protocordado são organismos exclusivamente marinhos. O registro fóssil indica que os urocordados surgiram durante o início do Cambriano por volta de 520 Mya (Chen et al., 2003).
Thaliacea
Thalicea é uma classe de pequenos animais marinhos planctônicos filtradores pertencentes ao subfilo Urochordata (Tunicados) que se alimentam essencialmente de plâncton. Estão descritas cerca de 1.250 espécies.
Com até 8 cm de comprimento, entretanto, apresentam tamanhos milimétricos, formam grande colonias gelatinosas e cada integrante são chamados de zoóides, agrupam-se geralmente em grandes colônias de consistencia gelatinosa que se move na coluna de água, embora em algumas fases do seu complexo ciclo de vida ocorram formas solitárias. São organismos pelágicos com distribuição natural nas regiões de águas quentes e temperadas de todos os oceanos e mares.
Conhecidos como taliáceos ou salpas, são animais marinhos pelágicos, de pequeno porte, normalmente com forma de "barril", com corpo gelatinoso e transparente, sobretudo as salpas e os doliólidos. O corpo é oco, recoberto por uma túnica complexa, com as vísceras a formar uma massa na superfície ventral. A musculatura é em bandas anelares distribuída por todo o corpo e não apenas nos sifões.
Apresentam uma faringe perfurada muito desenvolvida. O eixo oro-aboral é retilíneo e a cavidade faríngea e cloacal ocupa quase toda a parte axial do corpo, o que permite que o fluxo de água gerado seja utilizado para a alimentação, para o intercâmbio gasoso e para proporcionar ao animal (ou à colônia) propulsão. Assim, a água impulsionada pelas contração dos músculos anelares é usada na locomoção, respiração, excreção e alimentação.
Apesar da maioria dos Urochordata serem sésseis, os Thaliacea são filtradores de vida livre, pertencendo à comunidade planctônica, (holoplâncton = passam toda a vida fazendo parte do plâncton). Assim, ao contrário das ascídias, os membros deste grupo possuem vida livre durante todo o seu ciclo de vida. Apresentam formas solitárias ou em cadeias coloniais de vários metros, possuindo sifão inalante e exalante em posições opostas no corpo. Muitas espécies possuem órgãos luminosos, sendo comum a bioluminescência no grupo, com destaque para o gênero Pyrosoma.
Larvacea ou Appendicularia
São animais planctônicos presentes em todos os oceanos, principalmente em águas quentes costeiras de até 100 m de profundidade. Das cerca de 70 espécies descritas, 25 ocorrem no litoral brasileiro. Os membros da classe Larvacea vivem dentro de uma "casa" constituída de polimucossacarídeos secretados pelos indivíduos dessa classe. Eles mantêm as características de larva na vida adulta. Possuem notocorda, fendas faringeanas e cauda pós-anal, mas não se desenvolvem e permanecem com morfologia larval durante toda a vida. São organismos planctônicos que filtram partículas de plâncton através de uma rede situada nessa "casa" de polimucossacarídeos. Recentemente descreveu-se essa restrutura mucosa secretada ao redor do corpo do animal, e é bem maior do que se pensava anteriormente.


Relacionamento filogenético entre os membros dos cordados

Cladograma mostrando as plesiomorfia e sinapomorfia para entre os cordados
Conhecimento atual da filogenia de deuterostômios, com sinapomorfias para cada nó indicado na filogenia. Equinodermos e hemicordados são grupos irmãos, enquanto as relações de tunicados ainda não estão bem resolvidas. A localização da Xenoturbella ainda não está exatamente correta. Os caracteres cordados indicados na filogenia que unem tunicados com cefalocordados e vertebrados estão presentes apenas na larva semelhante a um girino, dos tunicados. (Modificado de Swalla 2001 e Zeng e Swalla 2005).
CLASSE ASCIDIACEA


Classe Ascidiacea
CLASSE THALIACEA



Classe Thaliacea
Um peixinho se "escondendo" dentro de um urocordado
CLASSE LARVACEA OU APPENDICULARIA

Classe Larvacea





Os Larvacea ou Appendicularia criam uma "corrente de alimentação" para filtrar as partículas de alimento (plancton) da água circundante. As setas pontilhadas indicam a direção da corrente através da "casa" (túnica).
Reconstrução 3D de um Urocordado Larvacea e a estrutura complexa de seu filtro interior. Fonte: Video by Monterey Bay Aquarium Research Institute and the Digital Life Projection/University of MassachusettsCredit

ASCIDIACEA
CARACTERÍSTICAS GERAIS
Tunicados ou urocordados (apendiculares, salpas e ascídias), cefalocordados (lanceletes) e vertebrados constituem os três grupos existentes de animais cordados. Tradicionalmente, os cefalocordados são considerados os parentes vivos mais próximos dos vertebrados, enquanto os tunicados devem representar a linhagem cordada mais antiga.
No entanto, as análises filogenéticas de dados genômicos indicam uma forte afinidade entre tunicados e vertebrados, enquanto os cefalocordados deveriam estar mais distantemente relacionados (Delsuc et al., 2006).
Os tunicados (urocordados) são os únicos animais conhecidos que tem a habilidade de produzir um polissacarídeo semelhante a celulose (altamente cristalino). O nome “Tunicata” é derivado do tecido tegumentar único denominado “túnica”, que contém as microfibrilas de celulose, secretada pela epiderme. Até o momento, microfibrilas I de celulose foram encontradas em quase todas as ascídias e taliáceos. Assim, a composição celulósica da túnica é considerada uma característica comum às ascídias e taliáceos (animais pertencentes ao subfilo Tunicata = urocordados).
Os urocordados ou tunicados podem ser fixos, de vida livre ou parasitários. Possuem notocorda na região da cauda, apenas na fase larvar.
Receberam o nome de tunicados por possuírem uma estrutura em forma de túnica, recobrindo todo o corpo do animal; daí, o nome do subfilo. A túnica exoesquelética é complexa, composta essencialmente por tunicina, um isômero da celulose. Fato esse que os distancia dos cefalocordados e cordados.
Apresentam uma faringe perfurada altamente desenvolvida; entretanto, no processo de metamorfose, o tubo nervoso dorsal oco é perdido, assim como a notocorda. Os tunicados são comuns nas regiões costeiras de todos os continentes. A maioria das espécies tem habito séssil; entretanto, os representantes das classes Larvacea e Thaliacea estão adaptados a uma vida planctônica.
Os urocordados são animais, em sua maioria sésseis, que vivem grudados a rochas submersas em costões rochosos, a algas de grande porte ou a outros substratos submersos.
A larva é livre natante, e apresenta as características básicas do clado dos cordados. Todavia, após a fase larvar e ao se fixar no substrato a larva passa por uma metamorfose onde a notocorda desaparece e o tubo neural dorsal regride. Nos indivíduos adultos, a única característica que ainda permanece e que lembra um cordado é a presença de fendas faringianas.

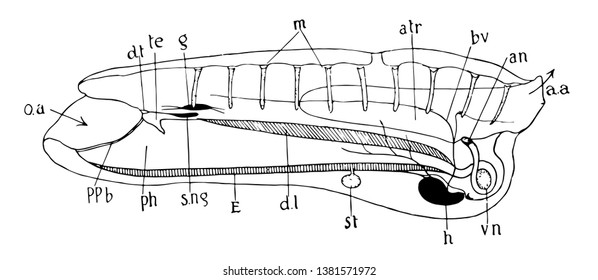
Anatomia de uma ascidiacea
Os urocordados alimentam-se de partículas orgânicas que são filtradas da água do mar que circula em seu corpo. O alimento adere ao muco produzido em um sulco da faringe chamado endóstilo, e é recolhido com auxilio dos cílios de células ciliadas e levado por estes cílios em direção ao esôfago e estômago, onde inicia a digestão extracelular. Do estômago o alimento passa para o intestino, onde a digestão termina extracelularmente e os nutrientes são absorvidos. Portanto são micrófagos filtradores.
Os resíduos são eliminados pelo ânus que se abre no sifão exalante, através do qual as fezes são eliminadas para o exterior junto com a água que vem das fendas branquiais.

Trocas gasosas
Nos urocordados a água que circula continuamente pelos sifões traz gás oxigênio e leva gás carbônico e excreções. As traves que separam as fendas faringianas apresentam uma rede de vasos capilares, sendo funcionalmente equivalente a brânquias. As trocas gasosas ocorrem na faringe, que recebe água continuamente. Esta possui uma grande superfície de contato com a água, realizando a troca de gás carbônico do sangue com o oxigênio da água. Os gases são transportados em solução física no plasma. Dentro da túnica há um espaço entre o organismo e a epiderme do manto, sendo essa também uma superfície de contato, em que ocorrem trocas gasosas cutâneas.
Sistema circulatório
O sistema circulatório é parcialmente aberto. A hemolinfa penetra em grandes bolsas (sinusóides) localizadas entre os tecidos corporais, e onde ocorrem as trocas gasosas entre o sangue e os tecidos. Os sinusóides são um tipo mais complexo de hemocela.
Os urocordados possuem um coração localizado na base da faringe do qual partem vasos que chegam às fendas faringianas (brânquias) e aos órgãos corporais.
Um fenômeno curioso que é típico dos urocordados é a reversão sistemática do fluxo circulatório. Após certo número de contrações do coração , em que a hemolinfa é impulsionada em direção às fendas branquiais faringianas, o coração para de se contrair por alguns instantes e ao reiniciar as contrações, impulsiona a hemolinfa em sentido inverso ou seja em direção aos órgãos do corpo.
Nesses animais, portanto, o sistema circulatório é composto por um coração ventral e vasos, que permitem um fluxo bidirecional de sangue, sendo único em todo o reino animal.
O sistema nervoso resume-se a receptores tácteis, e um “olho” (células fotorreceptoras) que percebe a variação da luminosidade.
Nesses animais, o sistema vascular, é bem desenvolvido e peculiar. Existe um coração em formato cilíndrico curvo ou em forma de U; dele partem dois vasos: um dorsal, que leva o sangue para alguns vasos menores da faringe (nos quais ocorrem as trocas gasosas), e outro ventral, que traz o sangue de volta para o seio venoso, passando, primeiramente, próximo ao intestino, no qual os nutrientes ficam retidos. O sangue apresenta corpúsculos que, embora não exerçam a função de transportadores de gases, realizam outros variados papéis. Dentre esses corpúsculos, podemos citar as células sangüíneas especializadas, os linfócitos, os quais originam outras células do sangue; os amebócitos que, nessa classe, têm função de carregar nutrientes, além de fagocitar resíduos não aproveitados; as células morulares, que concentram metais pesados, tais como o ferro (z:26, m:55,845), o nióbio (z:41, m:92,9) o tantálio (z:73, m:180,94) e o vanádio, que se localizam dentro de vesículas intracelulares. No caso do vanádio, acredita-se que seja usado na produção da túnica, devido ao seu poder redutor, e ainda pode ser usado como antibiótico, devido ao fato de a sua toxicidade desencorajar a predação aos tunicados.
A reversão periódica do fluxo sangüíneo, é uma característica única e peculiar dos tunicados, nos quais, a cada 2 ou 3 minutos, o batimento cardíaco cessa, sendo retomado na direção oposta. O coração é controlado por dois centros miogênicos (que fazem o coração bater), cada um localizado num ponto do órgão, sendo capazes de reverter a direção da circulação sanguínea pelo ponto sobre o qual houver maior pressão.
Sistema nervoso
Nos adultos, é relativamente simples, consistindo em um gânglio cerebral localizado entre os dois sifões. Tanto na cavidade visceral, como na cavidade branquial, observam-se neurônios que chegam às vísceras. Esses estão acoplados, superiormente, ao gânglio, no qual há uma glândula neural que produz neurohormônios importantes na reprodução; essa se abre na faringe, por meio de um tubérculo dorsal.
As larvas nadadoras (também conhecidas como livre-natantes) exibem tubo nervoso dorsal oco e uma pequena formação ganglionar.
Esses animais não apresentam órgãos sensoriais especiais; 9 entretanto, células de estimulação táctil (mecanossensoriais), localizadas entre o organismo e a túnica, são abundantes, provavelmente exercendo papel de controle da corrente de água que passa através da faringe
Reprodução
A reprodução pode ser assexuada por brotamento, e sexuada, e a maioria delas é monoica (hermafrodita).

Brotamento em ascidiacea




Reprodução sexuada e assexuada em Botryllus schlosserrina ascidiacea colonial .
O tunicado colonial Botryllus schlosseri é uma ascidiacea que vive em águas rasas e é facilmente criada em aquários. Seu peculiar ciclo blastogênico, caracterizado pela presença de três gerações blastogênicas (filtradores adultos, brotos e brotinhos) e por mudanças recorrentes de geração. Mais de 60 anos de estudos buscam compreender como a reprodução sexual e assexuada são coordenadas e reguladas no colônia. Esses animais possibilitam a utilização de diferentes abordagens metodológicas, desde a genética clássica até o transplante de células, contribuiu para o desenvolvimento desta espécie como um valioso organismo modelo para o estudo de uma variedade de processos biológicos. Dados dos estudos com essa espécie possibilitam um melhor entendimento da evolução dos cordados.
A ausência de estruturas excretoras limita a regulação osmótica, restringindo os urocordados ao ambiente marinho ou ligeiramente salobro. Sendo hermafroditas, os tunicados apresentam ciclos de vida complexos, incluindo alternância de gerações e formação de colônias; suas larvas assemelham-se às de girinos, e após uma existência livre natante se metamorfoseiam em animais adultos.
Ao passar da forma larvar para o estágio adulto, a cauda regride; com ela desaparecem a notocorda e grande parte do tubo neural (sistema nervoso). A larva dos tunicados é muito semelhante ao anfioxo (um cefalocordado) e à larva das lampreias (vertebrados bastante simples), o que evidencia o parentesco dos protocordados com os vertebrados. e também sugere que a evolução seguiu através de neotenia.
(Neotenia, é uma forma de pedomorfose, é a propriedade, em animais, de retenção, na idade adulta, de características típicas da sua forma jovem ou larval.(Neotenia é a retenção de características juvenis na forma adulta (maturidade)). Em linhas gerais, na neotenia os animais têm seu sistema reprodutor maturado e se reproduzem normalmente, porém seu aspecto externo, como um todo, é o de um indivíduo jovem. A maturação do sistema gamético difere daquela do sistema somático, o qual é reprimido. Neotenia é uma forma de pedomorfose. Em biologia do desenvolvimento, pedomorfose é um dos processos evolutivos aceitos atualmente para o surgimento de novas espécies e se refere ao evento no qual uma espécie descendente se reproduz em uma fase morfológica que era juvenil em seus ancestrais. Apenas mais recentemente esse processo de desenvolvimento corporal pôde ser extrapolado e entendido como um poderoso processo de formação de novos planos corporais.
Esse avanço no pensamento científico só veio a se formar quando a teoria da recapitulação de Ernst Haeckel, baseada na evolução por adição terminal, foi superada. Gould escreveu também sobre evolução por adição terminal, processo no qual a evolução ocorre com o acréscimo de um novo estágio no final do desenvolvimento. Um organismo primitivo teria parado em um estágio A e pela evolução surge um segundo estágio B, posteriormente um outro estágio C (A->B->C), e assim por diante. A cauda dos peixes em estágios evolutivos mais avançados como o linguado (Pleuronectes) pode ser usada como exemplo, ela passa por três etapas até chegar no estágio adulto. Essas fases se assemelham a estruturas presentes em peixes adultos de outras espécies que estariam mais próximas de peixes mais primitivos. Na fase larval (dificerca) a cauda seria parecida com a de um peixe pulmonado mais semelhante a um peixe primitivo, a segunda fase (heterocerca) teria a cauda próxima da do esturjão, e a terceira (homocerca) próxima da de um salmão, que seria a forma evoluída mais recentemente.
Esse processo de evolução resulta na recapitulação, mas não é a única forma que ela ocorre. Podem acontecer modificações em estágios mais iniciais do desenvolvimento e não necessariamente no final. A pedomorfose é uma heterocronia que pode ocorrer de duas formas; o tempo absoluto de desenvolvimento somático diminui enquanto o reprodutivo continua o mesmo (neotenia), ou o desenvolvimento reprodutivo acelera enquanto o somático permanece o mesmo (progênese). Nesse processo , em que a espécie evolui para reproduzir em uma fase precoce em relação ao ancestral, não ocorre a recapitulação completamente sua ancestralidade.
Na conclusão de seu livro Gould defende que nós humanos evoluímos através de um abrandamento do desenvolvimento, e mantemos muitas características que agora podem ser vistas nos estágios juvenis de nossos parentes primatas, e que sem dúvida caracterizam estágios dos nossos antepassados recentes. A Teoria da Recapitulação, ou Lei da recapitulação ontofilogenética, é mais conhecida pela expressão "A Ontogenia recapitula a filogenia" em que ontogenia refere-se ao desenvolvimento dos embriões de uma dada espécie e a filogenia refere-se à história evolucionária das espécies. A teoria faz uma relação entre evolução e desenvolvimento defendendo que o desenvolvimento do embrião repete o desenvolvimento evolucionário da espécie à qual pertence passando por etapas que se assemelham aos seus ancestrais na fase adulta. Em 1965, George Gaylord Simpson chegou a afirmar em "An Introduction to Biology" que "o fato de que a ontogenia não repete a filogenia está agora bem estabelecido”. Observações conectando evolução e desenvolvimento embrionário já haviam sido feitas por outros cientistas antes da teoria se publicada por Ernst Haeckel em 1866. Pessoas como Johann Friedrich Theodor Müller (naturalista teuto-brasileiro) e Edward Drinker Cope (Paleontólogo USA), reconheceram a recapitulação como uma chave para a reconstrução da filogenia.
Müller, observando um camarão marinho do gênero Penaeus, notou que ao nascer ele passava por uma fase de náupilo para depois chegar a fase de zoea. Com base nisso propôs que os caranguejos marinhos e os camarões que já nascem na forma de zoea, deveriam passar pelo estágio mais simples de náuplio durante seu desenvolvimento embrionário, o que se confirmou mais tarde. Diante deste fato alguns cientistas defendem que a Lei biogenética foi proposta inicialmente por Müller e copiada por Haeckel.
Charles Darwin obsevou que os embriões de vertebrados eram mais parecidos do que os vertebrados adultos, e que quanto mais jovem fosse o embrião, maior seria a semelhança. Erns Haeckel examinou com detalhes a interpretação de Darwin e propôs a teoria que designou como Lei biogenética (“Generelle Morphologie der Organismen” 1866). A lei foi criticada por muitos embriologistas da época alegando que diferenças significativas eram ignoradas nas comparações de Haeckel e que muitas de suas ilustrações foram alteradas.)

Desenvolvimento comparativo do mesoderma cardiofaríngeo em vertebrados (camundongo) e Urocordados (tunicados) (Ciona sp).
a) Embrião esquemático de dez dias E10 e a cabeça do camundongo no E14, e a árvore de linhagem que descreve as origens dos compartimentos cardíacos e músculos branquioméricos Em ratos. O primeiro campo do coração (FHF) e seus derivados são indicados em vermelho: ventrículo esquerdo (VE) e partes do átrio esquerdo (AE) e do átrio direito (AR); segundo campo do coração (SHF) e seus derivados estão em laranja: ventrículo direito (VD), partes dos átrios esquerdo e direito e trato de saída (OFT); os músculos esquelético branchiomérico estão amarelo; os músculos extra oculares estão em roxo. Todas as células derivam de progenitores pan-cardiofaríngeos comuns hipotéticos (verde escuro) que produzem o FHF e o segundo progenitor cardiofaríngeo Tbx1/10 + (CPM, verde claro). Linhas quebradas indicam que o FHF / SHF comum o progenitor ainda não foi identificado em camundongos.
b) Esquema dos diferentes tecidos musculares de Ciona juvenil, e a árvore de linhagens representando relações clonais e expressão gênica nos precursores cardiofaríngeos. Os primeiros precursores do coração (FHP) (vermelho) e os segundos precursores do coração (SHP) (laranja) contribuem para o coração (mistura de vermelho e laranja). As contribuições exatas do FHP e SHP para os compartimentos e tipos de células no coração juvenil ainda precisa ser elucidado. Os precursores musculares do sifão atrial (ASM, amarelo) formam o sifão atrial e os músculos longitudinais (LoM, amarelo) da parede do corpo. Os músculos do sifão oral (OSM, azul) surgem de uma linhagem não cardíaca diferente (A7.6, consulte o texto para obter detalhes). Células filhas do Mesp+. Os blastômeros B7.5 (brancos) produzem músculos anteriores da cauda (ATM, cinza) e células ventrais do tronco (TVC, verde escuro). Os últimos são progenitores pan-cardiofaríngeos que expressam Nk4 e se dividem assimetricamente para produzir o FHP (vermelho) e Tbx1/10+STVCs (disco verde claro). O último divide novamente assimetricamente para produzir SHP (laranja) e os precursores Ilhota+ de ASM e LoM. Figura adaptada de Diogo et alli. (Razy‑Krajka and Stol EvoDevo (2019) 10:13).
(Comparative development of cardiopharyngeal mesoderm in vertebrates (mouse) and tunicates (Ciona). a) Schematic of the mouse embryo at embryonic day (E) 10 and the mouse head at E14, and lineage tree depicting the origins of cardiac compartments and branchiomeric muscles in mice. First heart field (FHF) and its derivatives are indicated in red: left ventricle (LV), and parts of left atrium (LA) and right atrium (RA); second heart feld (SHF) and derivatives are in orange: right ventricle (RV), parts of left and right atria, and outfow tract (OFT); branchiomeric skeletal muscles are in yellow; extraocular muscles are in purple. All cells derive from hypothetical common pan-cardiopharyngeal progenitors (dark green) that produce the FHF and the second Tbx1/10+ cardiopharyngeal progenitors (CPM, light green). Broken lines indicate that the common FHF/SHF progenitor remains to be identifed in mice.
b) Schematic of the diferent muscle tissues of the Ciona juvenile, and lineage tree depicting clonal relationships and gene expression in the cardiopharyngeal precursors. The frst heart precursors (FHP) (red) and second heart precursors (SHP) (orange) contribute to the heart (red and orange mix). The exact contributions of the FHP and SHP to the compartments and cell types in the juvenile heart remain to be elucidated. Atrial siphon muscle precursors (ASM, yellow) form atrial siphon and longitudinal muscles (LoM, yellow) of the body wall. Oral siphon muscles (OSM, blue) arise from a diferent, non-cardiac lineage (A7.6, see text for details). Daughter cells of the Mesp+ B7.5 blastomeres (white) produce anterior tail muscles (ATM, gray) and trunk ventral cells (TVC, dark green). The latter are pan-cardiopharyngeal progenitors that express Nk4 and divide asymmetrically to produce the FHP (red) and Tbx1/10+STVCs (light green disk). The latter divide again asymmetrically to produce SHP (orange) and the Islet+ precursors of ASM and LoM. Figure adapted from Diogo et al. )
EVOLUÇÃO A PARTIR DE UROCORDADOS
Conhecendo agora as características dos protocordados, uma pergunta nos intriga: como ocorreu a evolução desses seres para vertebrados? Um pesquisador chamado Garstang, também intrigado com esse fato, propôs que os vertebrados se originaram de larvas semelhantes às de tunicados, as quais não sofreram metamorfose até a fase adulta (pedogênese), desenvolvendo gônadas funcionais e se reproduzindo. Uma outra hipótese, formulada por um pesquisador de nome Schaffer, postula que os vertebrados teriam se originado de larvas semelhantes às de cefalocordados, as quais, como na teoria de Garstang, não sofreram metamorfose até a fase adulta, desenvolvendo gônadas funcionais e se reproduzindo. Dessa maneira teriam surgido os primeiros vertebrados. Embora aceitas por diversos zoólogos, essas hipóteses permanecem sem provas.


Filo CHORDATA
Subfilo CEPHALOCHORDATA


Cefalocordados são um grupo de cerca de 25 espécies incluídas em três gêneros: Branchiostoma (peixes anfioxo ou lanceta), Asymmetron e Epigonichthys.
São exclusivamente marinhos e escavadores, embora possam nadar livremente na água, vivem enterrados na areia no fundo de mares rasos. Podem medir entre 5,0 e 8,0 cm de comprimento; carecem de pigmentos; são praticamente translúcidos. Seus músculos estão organizados forma de feixes segmentares (miômeros) em torno de 60.
Não apresentam uma cabeça (distinta) separada do tronco . Alimentam-se por filtração de de partículas orgânicas em suspensão.
Parede do corpo
Epiderme: Camada única; é ciliado no jovem e coberto com cutícula no adulto.
Sem glândulas ou cromatóforos.
Derme: é de natureza fibrosa, constituída de tecido conjuntivo,
Hipoderme (região subcutânea): é de natureza gelatinosa, mas também possui fibras. Musculatura: esplacnopleura produz na parte dorsal os segmentos musculares ou miótomos. Os miótomos são em forma de V, fibra estriada e separados por tabiques conjuntivos os mioseptos. Este arranjo é característico de todos cordados. Eles obliteram o celoma dorsal, deixando apenas uma pequena fração: miocele, reduzida a ranhuras e canais. Existem dois pequenos sacos celômicos.
Sistema digestório
Vestíbulo oral: suas bordas são prolongadas formando os cirros periorais, cerca de 30, de natureza sensorial. Abriga um conjunto de bandas ciliadas (órgão rotativo de Müller) que carrega partículas de comida para a boca; em seu centro se encontra a fosseta de Hatscher que é o orifício do primeiro saco celômico esquerdo.
Boca: ao redor dela há uma segunda coroa de tentáculos sensoriais (véu).
Faringe: Com cerca de 200 fendas branquiais; são direcionados obliquamente de
da parte dorsal para a ventral; entre eles está uma haste esquelética.
Elas abrem no cavidade peribranquial (átrio) através do atrióporo.
Em sua parte ventral está o endóstilo e em sua parte dorsal o sulco dorsal
epifaríngeo. Intestino médio ou mesodeu: a partir da frente um divertículo (ceco hepático) que se estende ao longo do lado direito da faringe.
A excreção é realizada pelos protonefrídeos ou solenócitos que estão localizados na região dorsal da faringe.
As trocas gasosas ou respiração é realizada principalmente pela pele por difusão direta (cutânea).
Sistema circulatório
O sistema circulatório é fechado e não apresenta um coração distinguível.
Apresenta uma série de vasos ventrais em que o o sangue circula para a frente e os vasos dorsais nos quais o sangue circula para a região posterior do corpo (para atrás). A circulação é realizada pela ação contrátil dos vasos que rodeiam a faringe. A circulação é lenta e a pressão do sangue é muito baixa. O sangue é incolor, pois não possui pigmentos respiratórios.
Retém todas as características sinapomórficas dos cordados ao longo da vida. Evidencia muito claramente as características diagnosticas dos cordados. Possuem também vários caracteres que sugerem um plano corporal vertebrado.
A notocorda, tubo nervoso dorsal oco, fendas branquiais e cauda pós-anal persistem em toda fase adulta. - Tem poucos centímetros de comprimento - Vivem com a parte posterior enterrada no sedimento e a parte anterior exposta para se alimentar.
Os anfioxos são filtradores, se alimentando pelo captura de particulado em redes de muco secretado ao longo das fendas faringeais. O batimento ciliar cria um fluxo de água para dentro da boca e para fora das fendas. Nos anfioxos, a faringe e as fendas faringeais são estruturas de alimentação e possuem papel minoritário na respiração, a qual primariamente ocorre ao longo da superfície externa do corpo, respiração cutânea.
O anfioxo frequentemente se desenterra e nada para outro ponto. Sua natação se assemelha a dos peixes, através de contrações coordenadas de blocos de músculos seriais (miômeros).A contração desses músculos em V flexionam a notocorda e produzem ondulações laterais que impulsiona o corpo para frente.
Os músculos segmentados desenvolvem-se de blocos do mesoderma, chamados somitos e arranjados serialmente ao longo de cada lado da notocorda do embrião.
Filo CHORDATA
Subfilo CRANIATA
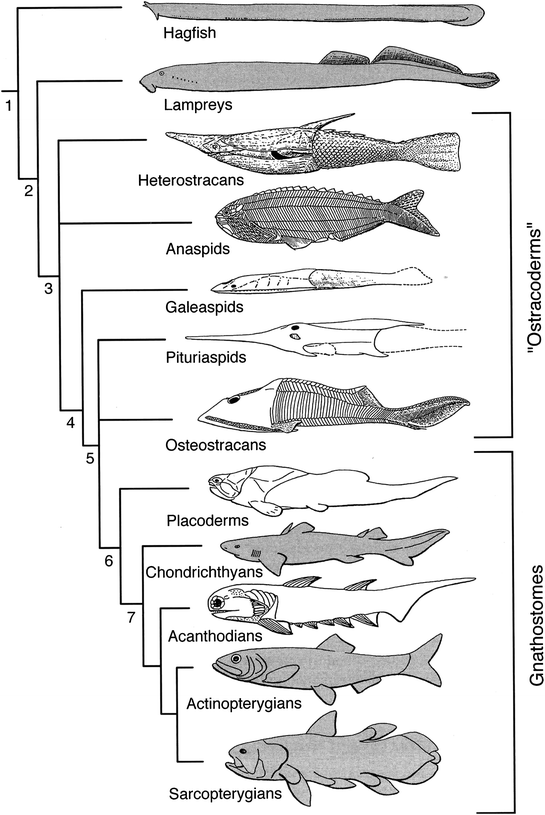
Superclasse AGNATHA OU CYCLOSTOMATA
Agnatha palavra criada para designar um grupo parafilético de animais cordados. Do grego: ἀ = a (negação) + γνάϑος = gnáthos, mandíbula, significa literalmente: animais sem mandíbulas. São cordados craniados, também chamados de ciclostomados com brânquias em forma de bolsa e sem mandíbulas, sem vértebras e com esqueleto cartilaginoso. Sugiram no período Ordoviciano 488 maa e chegaram ao presente.

Origem dos Agnatha ou Cyclostomata no ordoviciano

Cladograma mostrando a posição dos táxons
Cephalochordata, Urochordata e Craniata dentro dos cordados
São animais sem mandíbula. Dulcícolas e marinhos.
Possuem uma boca circular com dentículos de queratina e uma língua com dentes que usam para raspar o alimento. As lampreias podem perfuram a pele de peixes, golfinhos e tubarões dos quais se alimentam, sugando o sangue e tecidos musculares. São consideradas parasitas.
O corpo destes peixes (peixe-bruxa e lampreia) é longo e cilíndrico, com a parte caudal achatada lateralmente formando uma nadadeira.
Corpo revestido por pele fina, sem escamas.
Respiração branquial. As brânquias localizam-se dentro de bolsas branquiais, com abertura circular, na parede externa do corpo.
A notocorda constitui o único eixo de sustentação do corpo e persiste durante toda vida. Apresentam um crânio Crânio cartilaginoso.


Neurocrânio em larva de lampreia e peie bruxa
A excreção é feita por um par de rins, sendo a amônia e ureia o principal produto da excreção. Eles têm um sistema excretor que depende principalmente do trabalho dos rins. Em época de seca, Agnatha se enterra na lama e passa a produzir uréia. Os rins não apenas eliminam resíduos do corpo através do ânus, mas desempenham um papel fundamental na manutenção da regulação da quantidade de sal do corpo. Essas criaturas vivem tanto em água salgada quanto em água doce. Os rins devem trabalhar para manter um equilíbrio de sal quando em água doce. Quando o peixe está em água salgada, as brânquias do peixe trabalham para manter o excesso de sal fora.
As lampreias, devido ao seu hábito parasitário, têm sido uma praga séria de pescado Grandes Lagos da América do Norte, onde reduziram o número de trutas e outras espécies nativas. Por outro lado, os Agnatha têm pouca importância econômica. No Japão lampreias são usadas como alimento na preparação de molhos e caldos.
A superclasse Agnatha é composta de formas vivas e extintas.
As lampreias compartilham muitas semelhanças com os vertebrados com mandíbulas (gnatostomados), como a capacidade de regular a concentração de íons no corpo, a regulação nervosa dos batimentos cardíacos e olhos laterais bem desenvolvidos.
Os segmentos musculares, conhecidos como miômeros, consistem em fibras relativamente curtas que se inserem em septos de tecido conjuntivo, (os miocommata), entre os miômeros adjacentes.
Há apenas um esqueleto axial rudimentar e nenhum esqueleto apendicular.
Ordem MYXINIFORME
Uma família, dois gêneros, e 60 espécies.
Animais marinhos, altamente especializados em diversas áreas com na circulação, proteção (produção de muco) etc. Crânio cartilaginoso, não possuem vértebras e a sustentação do corpo fica a cargo apenas da notocorda que persiste por toda vida..
Nas Myxini o esqueleto axial é bastante incompleto (não mais que um cordão formado por nódulos cartilaginosos = notocorda), o que lhes permite enrolar o corpo num nó próximo a cauda que se desloca até a cabeça forçando a boca a arrancar um pedaço de carne da presa. O nó serve tanto para se libertarem de predadores, como para se alimentarem.
A pele é rica em glândulas mucosas, especialmente nas feiticeiras (Mixyne sp), que o produzem em grande quantidade para se defenderem de predadores.
O esqueleto é cartilaginoso, tal como os raios que sustentam as barbatanas dorsal e caudal em forma de remo. Não possuem nadadeiras pares.

Glândulas mucosas produzem muito e espessso muco
(Fonte: Angelena Mangieri)

Muco (Fonte BBC)
Ver vídeo nessa reportagem.
Artigo sobre a composição do muco

Apresentam pequenos tentáculos, com função sensorial, ao redor da boca.
Boca dotada de estruturas cartilaginosas móveis.
Dentículos que se projetam para a captura do alimento.
Alimentam-se de poliquetos e peixes vivos, mortos ou doentes, nos quais entram pelas brânquias ou ânus, comem os tecidos moles do interior do corpo da presa.
Pele com uma espessa camada de muco, para proteção. Muco com proteína capaz de formar fibras de grande resistência. Pesquisas mostram que essa proteína poderia ser utilizada para conter hemorragias em ferimentos.
São monoicos (apresentam sistema reprodutor feminino e sistema reprodutor masculino), mas apenas um sexo é funcional.

Myxini sp
Corpo alongado (formato de enguias).
Sem escamas.
Coloração rosácea a púrpura.
Adultos geralmente ultrapassam 1,0 m.
Inteiramente marinhos (Profundidades 25,0 -300,0 m).
Distribuição cosmopolita.
Associados a substratos lamacentos.
Alimentam-se de invertebrados bentônicos ou vertebrados moribundos.
Constroem galerias.
Vivem em colônias com razão sexual de até 100 fêmeas para cada macho.
Olhos pouco desenvolvidos e um par de canais semicirculares.
Coração com seio venoso, átrio e ventrículo, (corações acessórios)
Ausência de inervação cardíaca (modulados por hormônios).
Fluidos perfazendo mais que 10% do corpo.
Células sanguíneas c/ pouca afinidade por oxigênio.
Ausência de neuromastos na linha-lateral (embora apresentem fibras nervosas laterais). Neuromastos são células receptoras ciliadas juntamente com células acessórias. Estes são expostos através dos poros da linha lateral. É um órgão muito semelhante ao ouvido interno dos vertebrados. Percebem vibrações, variações de temperatura, ondas de choque e alguns parâmetros químicos na água.
Rins
Tecidos mineralizados ausentes (ossos, dentina e esmalte)

Boca de um peixe-bruxa
Sinapomorfias
Boca circular com língua cartilaginosa protraível
Duas fileiras de dentes córneos presentes na língua
Três a quatro pares de tentáculos ao redor da boca
Ducto nasofaríngeo
Série de glândulas ventro-laterais produtoras de muco
Ordem PETROMYZONTIFORME

Lampreia
As lampreias são agnatos de ambiente marinho ou dulcícola. Pode atingir até 1 metro de comprimento. Consideradas ectoparasitas porque utilizam sua boca em forma de ventosa para se fixar no corpo de outros peixes.
A boca da lampreia apresenta uma língua dotada de inúmeros dentículos de queratina no funil bucal, que utiliza para raspar a pele do hospedeiro até perfurá-la.
Glândulas salivares produzem anticoagulante. Alimenta-se de sangue e tecidos corporais das presas. Possui olhos grandes.

Comparação entre um cérebro de lampreia e o cérebro de um cação


Apresenta sete pares de fendas branquiais (laterais), a água entra pela boca e sai pelas fendas.
Esqueleto axial
Nas lampreias a notocorda persiste no adulto por toda vida, envolvida por arcos neurais imperfeitos, sendo o eixo de sustentação do corpo.
Na maturidade sexual, as espécies marinhas de lampreia vão para ambientes de água doce para se reproduzir. Fecundação ocorre externa.
As fêmeas colocam seus óvulos, cerca de 200 mil, em covas no fundo do rio, enquanto que os machos cobrem esses óvulos com seus espermatozoides.
Larva chamada de amocete, que é desprovida de olhos e dentes e que permanecerá enterrada na lama do rio, por até cinco anos. Alimentação por filtração. Após esse tempo enterradas, as larvas migram para os mares onde completarão sua metamorfose e onde ocorrerá o surgimento de olhos, boca e língua com dentículos.
Sistema circulatório
Coração com duas câmaras (átrio e ventrículo). Os peixes-bruxa apresentam ainda corações “acessórios” para aumentar o fluxo sanguíneo

Petromyzon sp. Seção longitudinal diagramática do coração.
As câmaras estão expostas, mostrando detalhes
internos (seg. Jollie)
As lampreias (Lampetra fluviatilis) são estenohialinas e fazem uma migração anádroma (do mar para água doce) no outono. Eles param de se alimentar durante a migração e morrem após a desova na primavera. Durante este período eles vivem em um ambiente hipotônico e enfrentam os problemas de hidratação excessiva e perda de eletrólitos. O último perigo é agravado pelo fato de que eles nunca se alimentam em água doce e, portanto, não podem obter sal de sua dieta, assim como fazem os teleósteos (peixes ósseos) de água doce. O papel dos rins e seu possível controle hormonal nesses processos não é apenas de interesse especial para a lampreia, mas também pode ser relevante para a compreensão da evolução da função renal em vertebrados.
Reprodução
A lampreia marinha tem comportamento anádromo e os adultos passam de 18 meses a dois anos se alimentando no mar antes de retornar para desovar em riachos de água doce. Quando a lampreia adulta voltar para água doce, seu sistema digestivo não funciona, as cápsulas de esmalte caem de seus dentes, eles
param de se alimentar e ficam cegos. A lampreia marinha adulta pode migrar centenas de quilômetros rio acima para encontrar um habitat adequado para a desova. Eles usam suas bocas especializadas para segurar-se em rochas úmidas ou outras estruturas para ajudá-los a superar obstáculos. Lampreias adultas marinhas não se alimentas em água doce (Flescher e Martini 2002).
A época de desova varia dentro do alcance da lampreia, mas em muitos locais do hemisfério norte, a desova ocorre do final de maio ao início do verão.
Ocorrem picos de desova quando água atinge temperaturas entre 17 a 19ºC (Applegate 1950; Beamish 1980). A Lampreia marinha constrói ninhos alongados de cascalho e pequenas pedras em corredeiras com 25 a 50 cm de profundidade.
Machos e fêmeas trabalham juntos na construção do ninho e mais de uma fêmea pode compartilhar esta atividade. Lampreias muitas vezes carregam pedras para o ninho com a boca e usam movimentos do corpo para criar um ninho livre de sedimentos que pode ter até 25 cm de profundidade e até 1,0 m de comprimento (Leim e Scott 1966; Scott e Scott 1988). Os ninhos podem permanecer visíveis por vários anos após a construção. Cada fêmea produzirá uma média de 230.000
(máximo 305.000) ovos adesivos, não flutuantes (Scott e Scott 1988; Leim e Scott 1966; Applegate 1950; Beamish 1980) e machos e fêmeas morrem após a desova. Os ovos eclodem após 10 a 13 dias (Piavis 1972) e as pequenas larvas movem-se rio abaixo para áreas de água parada de riachos e lagos (Wagner e Stauffer 1962), onde se enterram em substrato lamacento.
As larvas permanecem no substrato por quatro a oito anos ou até 10 anos (Scott e Scott 1988; Applegate 1950), filtrando a água e alimentando-se de algas e do plâncton a deriva. lampreias no estágio de larva não possuem dentes ou olhos, mas estão equipados com um capuz oral especializado para facilitar este estilo de vida (Flescher e Martini 2002). Normalmente em julho, as larvas das lampreias marinhas começam uma metamorfose que dura de quatro a seis meses. Durante a metamorfose, a lampreia larval perde o capuz oral e desenvolve dentes, olhos e rins. Quatro a seis meses depois, a lampreia recém transformada, emerge do substrato e inicia sua migração para o mar (100 a 200 mm de tamanho).
As lampreias marinhas adultas raspam com sua língua produzindo um buraco na lateral externa da parede corpo de sua presa para se alimentar de fluidos e tecidos. Se você puxar uma lampreia que se alimenta, você encontrará um grande buraco sob o local onde o disco bucal da lampreia estava localizado.

Sistema excretor
A excreção é feita por um par de rins mesonéfricos, sendo a amônia e ureia o principal produto da excreção, e um par de tubos que levam o produto da excreção, principalmente amônia, para a cloaca.
Eles têm um sistema excretor formado por rins mesonéfricos. Em época de seca, As lampreias se enterram na lama e passam a produzir uréia. Os rins não apenas eliminam resíduos do corpo através do ânus, mas desempenham um papel fundamental na manutenção da regulação da quantidade de sal do corpo. Essas criaturas vivem tanto em água salgada quanto em água doce. Os rins devem trabalhar para manter um equilíbrio de sal quando em água doce. Quando o peixe está em água salgada, as brânquias do peixe trabalham para manter o excesso de sal fora.

As lampreias, devido ao seu hábito parasitário, têm sido uma praga séria de pescado Grandes Lagos da América do Norte, onde reduziram o número de trutas e outras espécies nativas. Por outro lado, os Agnatha têm pouca importância econômica. No Japão lampreias são usadas como alimento na preparação de molhos e caldos.

Disco oral de uma lampreia adulta mostrando a
dentição e outras estruturas associadas.
A linha pontilhada delineia os vários campos.
(AF = campo anterior; AR = fileiras anteriores; ER = linhas exo-laterais;
F = fímbria oral; FAR = primeira fileira anterior; FPR = primeira linha posterior;
I = lâmina infraoral; LC = circum-orais laterais ou endolaterais;
LF= campo lateral; LL = lâmina lingual longitudinal;
MG= marginais; PF = campo posterior; PR = linhas posteriores;
SO = lâmina supra-oral; TL = lâmina lingual transversal
(Seg. de Hubbs e Potter (1971)).
Sinapomorfias
Ventosa oral suportada por cartilagem anelar.
Dentículos córneos no funil e na língua protrusível.
Glândula anticoagulante.
Ducto naso-hipofisário.


Historia evolutiva das Opsinas em vertebrados.
Opsina é a parte proteíca da rodopsina. Esse grupo de proteínas, sensível à luz, via cromóforo da retina (ou uma variante) encontrada nas células fotorreceptoras da retina. Cinco grupos clássicos de opsinas estão envolvidos na visão, mediando a conversão de um fóton de luz em um sinal eletroquímico, o primeiro passo na cascata de transdução visual. Outra opsina encontrada na retina de mamíferos, a melanopsina, está envolvida nos ritmos circadianos e no reflexo pupilar, mas não na visão. Opsinas também tem um papel no paladar ou em qualquer forma de sensibilidade química.

The retinal molecule inside an opsin protein absorbs a photon of light. Absorption of the photon causes retinal to change from its 11-cis-retinal isomer into its all-trans-retinal isomer. This change in shape of retinal pushes against the outer opsin protein to begin a signal cascade, which may eventually result in chemical signaling being sent to the brain as visual perception. The retinal is re-loaded by the body so that signaling can happen again.
(Fonte:OpenStax College 1, 2, 3 tipos de Opsinas)
1. Fendas branquiais externas
Aberturas que conduzem às brânquias internas usadas para extrair o oxigênio da água. As lampreias têm sete fendas branquiais circulais distintas.
2. Funil bucal
É o início da cavidade bucal. Contém os numerosos dentes da lampreia adulta. É circundado e apoiado pelo disco oral.
3. Papilas bucais
Projeções em forma de dedo que circundam o funil bucal.
4. Sistema de linha lateral
Um sistema que consiste em linhas de poros que detectam as correntes de água, mudanças de pressão da água e movimentos e vibrações na água. Os poros externos visíveis do sistema da linha lateral levam a um canal interno, que se conecta às células sensoriais especializadas. Acredita-se que esse sistema esteja relacionado ao sentido da audição em outros vertebrados.
5. Narina Medial
A narina medial é uma característica ancestral exclusiva das lampreias. Outros vertebrados têm narinas pares. A narina é responsável por detectar odores e leva a um tubo nasal na região dorsal da cabeça. Uma lampreia pode “cheirar” ao perceber produtos químicos na água. Essas partículas de cheiro podem ser detectadas a grandes distâncias. Aromas químicos entram na narina medial e passam pelo tubo nasal, ativando as células nervosas olfatórias ao longo do caminho, e enviam sinais elétricos ao cérebro através do nervo olfatório.
6. Órgão pineal
O órgão pineal está localizado sob a pele imediatamente posterior à narina medial. A evidência de sua presença é mostrada por uma saliência redonda generalizada no lado dorsal da cabeça. Na lampreia, o órgão pineal é às vezes chamado de “terceiro olho” devido ao seu papel na percepção da luz e da escuridão. O órgão pineal contém uma retina sensível à luz que não forma imagens visuais.
7. Olhos
Os olhos são órgãos sensoriais responsáveis por receber informações visuais. A recepção da imagem é feita por células especializadas (comes e bastontes) que transferem esse impulso ao nervo óptico, que envia esses impulsos visuais ao cérebro. No cérebro as imagens são decifradas e interpretadas. O olho da lampreia adulta é estruturalmente muito semelhante aos olhos de outros vertebrados, consistindo em córnea, íris, cristalino e retina. Não há pálpebras na lampreia.
8. Barbatana dorsal anterior e barbatana dorsal posterior
Ambas barbatanas são usadas para manter a orientação vertical na água enquanto o animal se desloca.
9. Barbatana Caudal
Uma barbatana poderosa usada para empurrar o corpo da lampreia na água.
10. Cloaca
A abertura comum dos sistemas urinário e reprodutivo. Recebe resíduos dos rins e fluidos dos órgãos reprodutores e os transfere para o meio externo pela abertura da cloaca. A papila urogenital é uma saliência que pode se estender da cloaca.
11. Ânus
O ânus está localizado imediatamente anterior à cloaca. É uma extensão do intestino por meio da qual os resíduos sólidos (fezes) são expelidos do corpo.

Tabela relacionando as diferenças entre peixe bruxa e lampreia
CRANIATA
(VERTEBRATA)
GNATHOSTOMATA
CHONDRICTHYES & OSTEICHTHYES


Vértebras torácicas
(De Anatomography em Anatomography (setting page of this image)
Cervical spine: 7 vertebrae (C1–C7)
Thoracic spine: 12 vertebrae (T1–T12)
Lumbar spine: 5 vertebrae (L1–L5)
Sacrum: 5 (fused) vertebrae (S1–S5)
Coccyx: 4 (3–5) (fused) vertebrae (Tailbone)





Filo CHORDATA
Subfilo Urochordata
Subfilo Cefalochordata
Subfilo Craniata
VÍDEOS
https://youtu.be/djUilMQleD8
Equinodermos (0 min-12min) e
Cordados (12min em diante)
CHORDATA
https://youtu.be/eUlM4rV9xl8
Características do filo Chordata
https://youtu.be/vQzUDrsWDvU
FILO: CHORDATA
https://youtu.be/9B547lihj6g
urocordados
EXERCÍCIOS
1. Alguns chefs de cozinha sugerem que o peru não deve ser preparado inteiro, pois a carne do peito e a da coxa têm características diferentes, que exigem preparos diferentes. A carne do peito é branca e macia, e pode ressecar dependendo do modo como é preparada. A carne da coxa, mais escura, é mais densa e suculenta e deve ser preparada separadamente. Embora os perus comercializados em supermercados venham de criações em confinamento, o que pode alterar o desenvolvimento da musculatura, eles ainda mantêm as características das populações selvagens, nas quais a textura e a coloração da carne do peito e da coxa decorrem da composição de suas fibras musculares e da adequação dessas musculaturas às funções que exercem. Considerando as funções desses músculos nessas aves, é correto afirmar que a carne:
a) Do peito é formada por fibras musculares de contração lenta, pobres em mitocôndrias e em mioglobina, e eficientes na realização de esforço moderado e prolongado.
b) Do peito é rica em fibras musculares de contração rápida, ricas em mitocôndrias e em mioglobina, e eficientes na realização de esforço intenso de curta duração.
c) Da coxa é formada por fibras musculares de contração lenta, ricas em mitocôndrias e em mioglobina, e eficientes na realização de esforço moderado e prolongado.
d) Da coxa é formada por fibras musculares de contração rápida, pobres em mitocôndrias e em mioglobina, e eficientes na realização de esforço intenso de curta duração.
e) Do peito é rica em fibras musculares de contração lenta, ricas em mitocôndrias e em mioglobina, e eficientes na realização de esforço moderado e prolongado.

2. Complete com os nomes e funções das estruturas dentro dos retangulos
3. Nos grupos de invertebrados estudados anteriormente, as características morfológicas sempre foram definidas a partir do estudo do animal adulto; entretanto, para os Chordata (protocordados e vertebrados), essa caracterização deve ser procurada na fase embrionária. Explique.
4. Os protocordados são os animais que mais se assemelham aos vertebrados do ponto de vista estrutural e do desenvolvimento. Com que argumentos você defenderia tal afirmativa?
5. A presença das fendas faríngeas é uma das características que aproxima os protocordados dos vertebrados. Discuta o papel funcional dessa estrutura para os protocordados do filo Urochordata.
5. A notocorda é uma estrutura muito importante. Na sua opinião, qual a melhor definição para notocorda e qual a sua importância?
http://www.biologydiscussion.com/animals-2/phylum-chordata/subphylum-vertebrate-craniata-phylum-chordata/40601
https://www.researchgate.net/publication/274398503_The_notochord_Structure_and_functions
https://go.gale.com/ps/anonymous?p=AONE&sw=w&issn=00368075&v=2.1&it=r&id=GALE%7CA99118703&sid=googleScholar&linkaccess=fulltext
{kind=link}
(ppt sobre órgãos dos sentidos)
Anatonia das lempreias
Myxiniformes
Evolução
Nam et ipsa scientia potestas est
(E portanto, conhecimento em si é poder)
Sir Francis Bacon.
Nenhum comentário:
Postar um comentário