Euplectella aspergillumé uma esponja de vidro do oceano profundo;
vista aqui a uma profundidade de 2.572 metros (8,438 ft) coasta da California, EUA.
Biodiversidade e morfotipos de esponjas na borda de uma parede marinha a 20 m de profundidade. Na foto está presente a esponja tubular amarela, Aplysina fistularis, a esponja de vaso roxa, Niphates digitalis, a esponja incrustante vermelha, Spirastrella coccinea, e a esponja corda cinza, Callyspongia sp.
Os
poríferos, filo Porifera: do latim porus = poro + ferre = portar; também conhecidos e chamados de espongiários, são animais aquáticos,
encontrados tanto em ambientes marinhos quanto de água doce.
São organismos conhecidos e utilizados desde a antiguidade, foram incluídas já no primeiro tratado de classificação de organismos, escrito em 350 a. C. por Aristóteles.
Preparando o esqueleto de uma esponja para banho
https://youtu.be/m8a0oNsDEx8
Alguns tipos de esponjas marinhas
Considerados inicialmente como plantas devido ao seu habito séssil e assimetria de muitos de seus representantes, sua natureza animal só foi reconhecida no final do século XVIII em 1765 quando se observou a corrente de água em seu corpo. Durante
grande parte do século XIX Porífera e Cnidaria eram colocados juntos no táxon
Radiata ou em Coelenterata (Santos, 2016).
Foi o naturalista inglês R.E. Grant quem primeiro
compreendeu a anatomia e fisiologia das esponjas e criou o nome Porífera, que
significa “portador de poros".
As esponjas são os animais mais
simples dentre os Metazoários, apresentando uma organização morfológica bastante diferente
da dos outros animais.
Devido a esta característica muito peculiar, pesquisadores resolveram considerá-las um grupo a parte dentro do Reino Animalia ou Metazoa (Santos, 2016).
Segundo Hajdu et al. (2011), tanto dados moleculares como morfológicos sustentam a
inclusão dos Porifera em Metazoa.
Dentre os aspectos mais importantes podemos listar cinco características compartilhados pelas esponjas e os demais metazoários:
1) multicelularidade diplóide,
2) meiose,
3) padrão da oogênese,
4) padrão da espermatogênese,
5) plano básico da célula do esperma
6) plano básico das células flageladas: opistoconta.
Esponjas são animais bentônicos sésseis filtradores de partículas em suspensão, que utilizam células flageladas chamadas coanócitos para promover a circulação da água através de um sistema de canais exclusivo do filo, o sistema aquífero. Diferentemente de outros grupos de organismos bênticos e filtradores, por exemplo, moluscos, ascídias e certos poliquetas, que quase não retêm as partículas inferiores a 1 ou 2 μm, as esponjas utilizam matéria orgânica particulada de 0,1 a 50 μm e sua associação com algas cianofíceas e bactérias lhes permitem utilizar matéria orgânica dissolvida (VACELET, 1979, Santos, 2016).
Uma exceção a isso é encontrada em algumas esponjas carnívoras, altamente adaptadas às águas profundas, em que o sistema aquífero é inexistente, mas que têm uma superfície exterior pegajosa com a qual as pequenas presas são capturadas (VACELET; BOURY-ESNAULT, 1995, Santos, 2016).
Apesar da aparente simplicidade quando comparado a animais mais complexos, o padrão de organização das esponjas é indiscutivelmente eficiente, tendo garantido sua presença nos ecossistemas marinhos, e em habitats de água doce, por mais de meio bilhão de anos (HAJDU et al., 2004). Por ser considerado um grupo de animais mais antigo existente, as esponjas estão entre os animais mais primitivos (ERPENBECK; WÖRHEIDE, 2007). O primeiro registro fóssil data de 750 milhões de anos (MA), e seu bauplan é basicamente inalterado desde o Cambriano superior (509 MAA, REITNER; WÖRHEIDE, 2002, Santos, 2016).
Embora as esponjas sejam conhecidas da humanidade desde as civilizações mais antigas (~4.000 anos [VOULTSIADOU et al., 2011]), elas não foram reconhecidas como uma linhagem metazoa independente até meados do século XIX, quando Robert Grant (GRANT, 1836) observou pela primeira vez a morfologia e fisiologia única, e criou o nome Porifera para elas, embora outros nomes tenham sido frequentemente utilizados (e.g. “Spongida”, “Spongiae”, “Spongiaria”).
Desde então, a espongologia, o estudo de todos os aspectos da biologia, ecologia, taxonomia e química de esponjas, tornou-se uma disciplina atraindo uma população cada vez maior de centenas de pesquisadores em todo o mundo. Hoje vemos que cada vez mais, as esponjas são estudadas como parte de um empreendimento mais amplo de tentar detalhar a Árvore da Vida (VAN SOEST et al., 2012).
Características gerais
São organismos sésseis, por não possuírem estruturas locomotoras, serem pouco móveis (apenas a larva é livre natante) e viverem fixos ao fundo marinho ou a rochas em rios e lagos, solitários ou em colônias; alguns podem fazer parte de recifes. O tamanho, forma e coloração variam bastante, de acordo com a espécie, fatores ambientais e associações com algas unicelulares e bactérias fotossintetizantes.
Dysidea sp. com cianobactérias filamentosas longas vivendo em simbiose com a esponja em seu corpo (mesohílo), contribuindo para a nutrição da esponja por meio da fotossíntese.
As esponjas também são verdadeiros "hotéis" para uma miríade de outras formas de vida. Isso inclui camarões, cracas, vermes e, mais significativamente, os outros Domínios da Vida (Bactérias e Arquéias). Em muitas esponjas, geralmente há mais células de bactérias do que células de esponja, e algumas delas co-evoluíram com as esponjas. Esses parceiros coevolucionários incluem as cianobactérias que vivem no mesohilo. Essas bactérias vivem em simbiose com muitas esponjas, principalmente em águas rasas, fornecendo nutrientes e subprodutos fotossintéticos à esponja por meio da fotossíntese.
Apresentam corpo muito simples, sem tecidos diferenciados.
Apresentam poros por toda extensão (superfície) corporal formados por células denominadas porócitos; há também aberturas maiores chamados de óstios, estruturas que facilitam a comunicação, dos coanócitos (células que promovem o fluxo de água e a captura do alimento), e o átrio ou espongiocele, e dai para o ósculo onde a agua sai do corpo do animal.
Fluxo de água e camadas celulares
É pelos porócitos e óstios que a água entra no interior do animal, passando pela coanoderme ou camada de coanócitos, e onde é filtrada. Depois a água ganha a cavidade central a espongiocele ou átrio, e é expelida continuamente pelo ósculo.
O ósculo, na porção superior, é a maior abertura do corpo das esponjas e por onde a água sai, eliminando excretas e outros resíduos,
inclusive gás carbônico. Por isso as esponjas são consideradas animais
filtradores. São filtradores incanssáveis, para conseguir 30 gramas de alimento esses animais tem que bombear e filtrar uma tonelada de água.
Externamente as esponjas são revestidas com células especiais os pinacócitos, células achatadas e unidas que revestem e protegem.
Internamente, são os coanócitos:
células ovoides, dotadas de flagelos e colarinho (coana), responsáveis pela circulação da água no corpo da esponja. Os coanócitos são células responsáveis pela
captura de partículas alimentares, digestão intracelular (que ocorre no citoplasma), e a transferência dos nutrientes para os amebócitos, que são células localizadas no mesohilo ou mesênquima. Os amebócitos são responsáveis pelo transporte dos nutrientes para as demais células do corpo da esponja.
No mesohilo encontramos também, células encarregadas da regeneração celular e reprodução os arqueócitos e célulasresponsáveis pela secreção de estruturas de sustentação esquelética os escleroblastos que produzem as espículas, que são estruturas formadas de carbonato de cálcio ou sílica, ou fibras
proteicas, principalmente a espongina.
As esponjas podem ser monoicas ou dioicas. Reproduzem-se assexuadamente por
brotamento, fragmentação e, ainda, por gemulação, no caso das esponjas de água
doce.
Na reprodução sexuada os arqueócitos e coanócitos se modificam em
óvulos e espermatozoides, que são liberados para o ambiente externo pelo ósculo e para
dentro de outras esponjas, por meio das correntes de água.
Habitat
As esponjas são dominantes em costões rochosos, recifes de coral e em substratos artificiais (BERGQUIST, 1978), fazem parte também principalmente nos ambientes ciáfilos (pouca luminosidade), grutas submarinas e cavernas (VACELET, 1979; VAN SOEST, 2009). Atualmente as esponjas marinhas são encontradas em todos os oceanos, desde a zona do entre-marés até profundidades que chegam a mais de 8.800 metros. Nos ambientes profundos os poríferos são frequentemente dominantes, estando as maiores riquezas conhecidas associadas a locais de maior complexidade estrutural, usualmente encontrados em setores de declividade mais acentuada (talude, cânions, montes submarinos), onde os substratos são mais consolidados (HAJDU et al., 2011). Segundo este autor, nesses “oásis” profundos, os poríferos podem desempenhar um papel estruturador, conferindo maior complexidade tridimensional como fazem em ambientes rasos. São encontradas em ambientes dulcícolas sobretudo lóticas (rios e córregos), aderidas a rochas ou a galhos e troncos submersos ou flutuantes.
Embora quase toda a totalidade das esponjas seja marinha, há, em todo o mundo, mais de 200 espécies de poríferos de água doce registradas, sendo pouco mais de 50 a 54 espécies encontradas no Brasil.
Na ciência, é consenso que elas surgiram a partir de esponjas marinhas e migraram ou se adaptaram de alguma maneira ao ambiente dulcícola, embora ainda não se saiba se tal fato ocorreu várias vezes ou somente uma, durante o processo evolutivo.
Alguns poríferos de água doce desempenham relações ecológicas positivas com outros organismos. Um exemplo típico é o fato de atuarem como refúgio e/ou abrigo para diversos animais. Outro fator envolve a relação simbiótica entre elas e algas verdes unicelulares. Neste caso, tais esponjas são consideradas fototróficas, já que grande parte de sua nutrição é oriunda das atividades fotossintéticas das algas simbiontes (Araguaia,s/d).
O mais abrangente estudo
já realizado foi feito por Batista & VolkmerRibeiro (2002), no rio Manso, tributário do
rio Cuiabá, que contribuiu para o levantamento da fauna espongológica, revelando a
presença de seis espécies (Corvospongilla
seekli, Eunapius fragilis, Troehospongilla
repens, Oneosclera navieella, Oneosclera
petrieola, Drulia uruguayensis) (Marostega, et alii, 2013).
Estudos sobre tecidos animais em esponjas
Além de simplesmente colar e manter unidas as células, as moléculas de adesão celular são reguladas dinamicamente durante o desenvolvimento dos organismos e a migração celular, reguladas espacialmente em tecidos polarizados e envolvidas na sinalização celular e na mecanotransdução. Consequentemente, uma miríade de mecanismos de adesão evoluiu para funcionar em diferentes contextos em animais. Destes, dois predominam: (1) a junção aderente, que está envolvida na adesão célula-célula e é composta de receptores de caderina, α- e β-catenina, e (2) adesões focais, que estão envolvidas na adesão célula-matriz extracelular (ECM) (cell–extracellular matrix adhesion) e composta por proteínas, como integrinas, vinculina, paxilina, talina e quinase de adesão focal (FAK) (focal adhesion kinase)(Mitchell and Nichols, 2019).
Os componentes moleculares da junção aderente e das adesões focais são amplamente conservados em animais, e alguns de seus componentes têm origem fora dos animais (1*). No entanto, os estudos experimentais da composição e função das junções celulares são amplamente restritos a animais bilaterianos, como a lombriga Caenorhabditis elegans, a mosca da fruta Drosophila melanogaster e vertebrados. Estudos recentes demonstram papéis conservados para proteínas de junção aderentes também em cnidários.
Schematic representation of integrin-mediated cell-adhesion and cell-signaling evolution. Left: The canonical metazoan integrin adhesion complex. The colors correspond to the three main steps in the evolution of the integrin adhesion mechanism, as shown in the cladogram. Dots indicate origin, and crosses indicate losses. The branch leading to Amastigomonas sp. is shown dashed, because its phylogenetic position remains unresolved (see main text for discussion).
Schematic representation of the eukaryotic tree of life showing the distribution of the different components of the integrin adhesion complex. The number of integrin homologs is shown. A black dot indicates the presence of clear homologs, whereas a hollow dot indicates the presence of putative or degenerate homologs. Absence of a dot indicates that a homolog is lacking in that taxon. The phylogenetic relationships are based in several recent phylogenetic studies.
Portanto as esponjas (Porifera) são organismos de importância crítica para reconstruir as etapas iniciais na evolução dos mecanismos de adesão de células animais (Mitchell and Nichols, 2019).
Eles são um dos grupos de animais mais divergentes filogeneticamente, sua anatomia é fundamentalmente diferente de outros animais, e há questões de longa data sobre a estrutura e homologia de seus tecidos em comparação com epitélios em outros animais. Tem sido argumentado que a adesão celular em esponjas (classe Demospongiae, em particular) depende principalmente de um complexo de proteoglicano extracelular denominado Fator de Agregação. Consequentemente, acredita-se que a integridade dos tecidos de demospongie dependa da interação das células com o fator de agregação, um componente da ECM, em vez de por meio de junções celulares como as encontradas em epitélios de outros animais (Mitchell and Nichols, 2019).
A integridade e organização dos tecidos animais dependem de complexos proteicos especializados que medeiam a adesão entre as células (junções aderentes à base de caderina) e com a matriz extracelular (aderências focais à base de integrina). Reconstruir como e quando essas junções celulares evoluíram é fundamental para compreender a evolução inicial dos tecidos em animais.
Examinamos homólogos de proteínas de adesão focal em tecidos da esponja de água doce, Ephydatia muelleri (filo Porifera; classe Demospongiae).
As principais descobertas foram:
(1) que os homólogos de adesão focal da esponja (integrina, talina, quinase de adesão focal, etc.) co-precipitam como um complexo, separado das proteínas de junção aderentes;
(2) que estruturas à base de actina que se assemelham a aderências focais se formam na interface célula-substrato e sua abundância é regulada dinamicamente em resposta ao cisalhamento de fluido;
(3) as proteínas de adesão focal localizam-se nas adesões célula-célula e célula-matriz extracelular, e;
(4) a proteína de junção aderente β-catenina é co-distribuída com proteínas de adesão focal em junções célula-célula em todos os lugares, exceto na coanoderme, e em novas junções entre células com espículas e entre células com bactérias ambientais (and between cells with environmental bacteria). Esses resultados esclarecem a diversidade, distribuição e composição molecular das junções celulares em tecidos de E. muelleri.
Adherens Junction ou junções aderentes, e estruturas semelhantes à adesão focal em tecidos de Ephydatia muelleri. (a) Topo: as células da endopinacoderma apical contêm feixes de filamentos de actina que culminam em placas densas que lembram junções aderentes em pontos de contato célula-célula. Embaixo: as células da basopinacoderme contêm feixes de filamentos de actina que se assemelham a fibras de estresse de aderências focais (pontas de flechas brancas). (b) Diagrama transversal do corpo juvenil de E. muelleri ilustrando a distribuição de Emβ-catenina (as setas azuis indicam a direção do fluxo de água no sistema aquífero). (c) Ilustração esquemática da organização molecular de uma adesão focal. As proteínas destacadas em vermelho são o assunto do estudo atual [FAK = Focal Adhesion Kinase.(Mitchell and Nichols, 2019)
Adherens Junction- and Focal Adhesion-like structures in tissues of Ephydatia muelleri. a Top: Cells of the apical endopinacoderm contain bundles of actin filaments that culminate in dense plaques resembling spot adherens junctions at points of cell–cell contact. Bottom: Cells of the basopinacoderm contain bundles of actin filaments that resemble stress fibers of focal adhesions (white arrowheads). b Cross-sectional diagram of the juvenile E. muelleri body illustrating the distribution of Emβ-catenin (blue arrows indicate the direction of water flow in the aquiferous system). c Schematic illustration of the molecular organization of a focal adhesion. Proteins highlighted in red are the subject of the current study [FAK = Focal Adhesion Kinase. (Mitchell and Nichols, 2019).
Diferentes morfotipos de esponjas (asconoide, leuconoides e siconoides)
e células coanócitos e amebócitos.
ANATOMIA DE UMA ESPONJA
Saturnospongilla carvalhoi.
A) Espécime sobre pequeno graveto;
B, C, D) Gêmulas discóides em detalhe
(Foto U.S. Pinheiro)
Microscopia eletrônica de varredura de Saturnospongilla carvalhoi.
A) Megasclera; B-F) Gemoscleras; G, H) Gêmulas discóides
(Foto U.S. Pinheiro)
Microscopia eletrônica de varredura de Ephydatia sp.
A, B) Megascleras; C) Gemoscleras; D,E,F) Gêmula
(Foto U.S. Pinheiro)
Gêmula. Esquema mostrando uma gêmula de uma esponja de água doce.
Pode-se ver a micrópila, as espículas anfidiscos, as fibras de espongina entre as espículas e os arqueócitos que preenchem todo o espaço interno da gêmula. As gêmulas são resistentes a dessecação (ressecamento), congelamento, e anoxia (falta de oxigênio) e podem permanecer nesse ambiente por grandes períodos de tempo. Gêmulas são similares ao endósporo bacteriano (produzido para resistir a situações desfavoráveis). As espículas são constituidas de amebócitos circundados por uma camada de espículas e pode sobreviver a condições que matariam a esponja adulta. quando o ambiente tornar-se menos hostil a gêmula regenera todo o individuo.
Na figura acima são apresentados o eixo basal-apical e o polo anterior e posterior de algumas esponjas. (Eixo apical-basal e polo anterior-posterior das larvas e adultos de algumas espécies de esponjas). (a) MEV da larva de Clathriina reticulata (Calcinea) larva. (b) MEV da larva de Halisarca dujardini. (c) Microscopia ótica de Esperiopsis koltuni parenchymella. (d) Polymastia artica do Mar Branco (White Sea). (Courtesy of M. Fedjuk). (e) Sycon sp. White Sea (Courtesia of M. Fedjuk). (f) Haliclona aquaeductus do White Sea (Courtesy of M. Fedjuk). As flechas indicam a posterior–anterior (a–c) e o eixo basal–apical (d–f). ap: polo anterior, pp: polo posterior, o: ósculo.
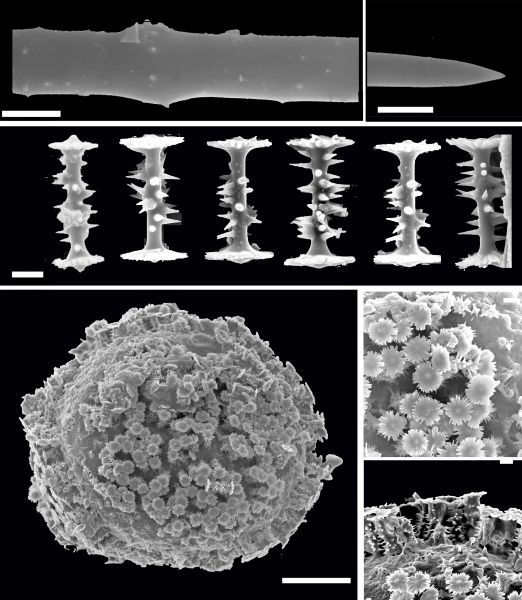
ESPÍCULAS DE ESPONJAS
Os elementos esqueléticos siliciosos das esponjas (filo: Porifera), denominados espículas, possuem várias características únicas que os distinguem dos elementos esqueléticos encontrados em outros Metazoa.
Eles são feitos de sílica [(SiO2)n] em vez de minerais à base de cálcio com uma precisão incomparável, dando origem a estruturas complexas espécie-específica. Essas estruturas geneticamente controladas e produzidas biologicamente são formadas em condições fisiológicas amenas, sem altas temperaturas, pressões ou produtos químicos cáusticos. As espículas são o determinante estrutural crítico que controla a morfologia das esponjas. No centro das espículas encontra-se um canal axial de 0,5–4,0 μm de largura que abriga o filamento axial orgânico. Desde sua descoberta, o filamento axial tem sido considerado um modelo que controla a morfologia das espículas. Um grande passo para o entendimento da morfogênese geneticamente controlada das esponjas foi a identificação da proteína estrutural das espículas, denominada silicateina, que está localizada tanto no filamento axial quanto na superfície das espículas. A silicateina é uma enzima que forma a bio-sílica necessária para a construção das espículas das esponjos. A formação das espículas é um processo rápido, que dura para uma espícula com comprimento de 190 μm e diâmetro de 6 a 8 μm a 21ºC apenas 40 horas. Por causa dessa alta taxa de crescimento, não foi claro por um longo tempo se a formação da espícula começa intra ou extracelularmente. Abaixo vemos esquematicamente a síntese de uma espícula simples e a sequencia dos passos principais no tempo e onde ocorre.
(A) A síntese da espícula (sp) começa intracelularmente nos esclerócitos (scl). As espículas primordiais estão associadas a filamentos (fi) que se presume que participam da extrusão da espícula em crescimento. Esta fase é dominada pela expressão de silicateina que, no estágio posterior, é necessária para a formação do núcleo e do cilindro da casca do manto silicioso da espícula. As moléculas de silicateina recém-formadas passam por uma organização fractal. (B) A espícula primordial é extrudada e torna-se associada no espaço extracelular com esclerócitos (scl) que intracelularmente formam os silicassomas (sis). Essas organelas contêm silicateina e silicato que são liberados no espaço extra-espicular e causam a formação de bio-sílica. (C) O crescimento da espícula (sp) continua em duas direções; alongamento axial e crescimento aposicional/espessamento (axial elongation and appositional growth/thickening). A formação da bio-sílica é mediada pela silicateina (sil) sob o consumo do substrato silicato (si). O crescimento da espícula é conduzido longitudinalmente e (subsequentemente) radialmente ao longo da protrusão celular. Durante esta fase, as extensões celulares se alongam por evaginação. O núcleo do manto da espícula é formado por silicateina, existente no canal axial, e a casca por silicateina, espalhada na superfície externa da espícula em crescimento. (D) Conclusão final do tamanho e forma da espícula. Após o término, a espícula se desconecta do esclerócito (não mostrado no esquema) e o orifício é fechado pela formação de biossílica. A direção do movimento da célula é indicada por uma seta (no interior dos retângulos) (Wang et alii, 2011).
Esponja marinha
TIPOS ESTRUTURAIS
Tipos estruturais presente nas esponjas:
Asconóide (A), Siconóide (B) e Leuconóide (C)
São reconhecidos três tipos estruturas de esponjas: ASCONÓIDE (A), SICONOIDE (B) e LEUCONÓIDE (C), que diferem entre si pela complexidade da parede do corpo e numero de coanócitos. O tipo asconóide é o mias simples dos três tipos. A parede é fina e possui poros inalantes que se abrem diretamente no átrio ou espongiocele. Este tipo é apresenta apenas uma camada coanócitos que reveste o átrio. Ex. gênero Leucosoleina. já nas esponjas do tipo siconóide, a parede do corpo é formada por projeções sinuosas em forma de dedos. Identificam-se dois tipos de canais: os inalantes e os radiais. A água penetra pelas camadas radiais, indo para a espongiocele. Os canais radiais são revestidos internamente por coanócitos. No tipo leuconóide, a parede do corpo é muito espessa e percorrida por um sistema de canais. Há canais inalantes e exalantes e, entre eles, câmaras revestidas por coanócitos. A água penetra pelos canais inalantes, passa por câmaras vibráteis (repletas de coanócitos) e vai à espongiocele através dos canais exalante.
Reino Metazoa
Sub reino Parazoa
Filo Porifera
Classes:
Calcarea
Demospongiae
Hexactinellida
Sclerospongiae
Diferentes aspectos da organização das esponjas, desde seu mutualismo com bactérias e algas, seu estágio anfiblástula, a forma de seus ancestrais hipotéticos e organização dos tipos Asconóide, Leuconóide e Siconóide, são apresentados.
Evolução da complexidade das câmaras flageladas dentro do filo.
Pinacócitos são células achatadas, irregulares, fortemente unidas que revestem e protegem todo o corpo da esponja.
PORÓCITOS
Porócitos são células dotadas de um poro central que permite a permite a passagem da água do meio externo para o átrio ou espongiocele.
COANÓCITOS
Coanócitos são células flageladas com uma expansão membranosa protéica em forma de colarinho (coana), que revestem a espongiocele e outras câmaras vibráteis internas das esponjas do tipo siconoides e leuconoides. O movimento dos seus flagelos cria a corrente de água que traz nutrientes e gases. Os nutrientes são filtrados pelo colarinho da célula, (estrutura gelatinosa semi-sólida, constituída antes por um conjunto de pequenos bastonetes eretos e separados por espaços. Qualquer partícula orgânica ou microrganismo planctônico (bacterias, algas, fungos, protozoários e resíduos) aprisionado no colarinho é encaminhado para baixo, em direção ao corpo celular onde ocorre a fagocitose. É no coanócito que ocorre a digestão intracelular, nos vacúolos digestivos que se ligam a lisossomos (organela que transporta enzimas digestivas).
MESÊNQUIMA OU MESOHILO
Mesênquina ou mesohilo é a região do corpo compreendida entre
a pinacoderme (camada protetora externa) e a camada de coanócitos (filtradora). É uma camada gelatinosa proteinácea
onde se encontram os amebócitos, os arqueócitos, os colêncitos os
escleroblastos e as espículas.
ARQUEÓCITOS
Arqueócitos são células indiferenciadas
totipotentes, que podem dar origem aos diversos tipos celulares. Têm papel
fundamental na distribuição de nutrientes e secundário na digestão dos
alimentos. Originam brotos e gêmulas, ambos estruturas reprodutivas, bem como
óvulos e espermatozóides.
AMEBÓCITOS
Amebócitos são célula totipotentes, com formato amebóide, que se movem por pseudopodes e que se
originam dos arqueócitos. Podem se diferenciar nos outros tipos de células, e tem papel importante no transporte de nutrientes entre os coanócitos e os outros tipos de células do corpo da esponja.
ARQUEÓCITOS
A totipotência refere-se à capacidade da maioria, senão de todas as células do corpo das esponjas, de mudarem de forma e função de acordo com as necessidades específicas do indivíduo. As células totipotentes (como as células-tronco) ocorrem em todo o reino animal, mas sua capacidade de mudança geralmente ocorre apenas em uma direção: de uma célula-tronco para outra célula derivada (especializada). Nas esponjas, essas mudanças podem ocorrer em ambas as direções. Existem centenas de tipos diferentes de células nas esponjas, muitas cujas funções ainda não são conhecidas.
Os mais primitivos são os Arqueócitos (células fagocíticas amebóides que são as principais células de transporte para alimentação, digestão, transporte de oxigênio e reparo do corpo). Nas esponjas, os arqueócitos podem se desenvolver em qualquer outro tipo de célula e, em seguida, voltar quando necessário à forma anterior. Outras células incluem células esféricas (com funções de excreção e defesa química), espongiócitos (produzem colágeno) e escelerócitos (secretam espículas), entre muitas outras. As esponjas têm um metabolismo intracelular: elas capturam, comem, digerem alimentos e excretam produtos residuais dentro das células, e não em qualquer cavidade corporal comum. A alta mobilidade das células é, portanto, crítica para a sobrevivência da esponja.
ESCLERÓCITOS OU ESCLEROBLASTOS:
Esclerócitos ou escleroblastos são células amebóides cuja função é
formar as espículas.
COLÊNCITOS
Colêncitos são células fixas que ficam ancoradas por fibras citoplasmáticas e são especializadas em secretar colágeno.
LOFÓCITOS
Lofócitos são células móveis, que
secretam as fibras de colágeno dispersas.
ESPONGIÓCITOS
Espongiócitos são células encarregadas da secreção das fibras de espongina, para formação do esqueleto das esponjas.
ESPONGIOBLASTOS originam as fibras de espongina (formada por colágeno)
ÁTRIO OU ESPONGIOCELE
Átrio ou espongiocele é o espaço ou cavidade interna nas esponjas asconóides. A parede do corpo das esponjas delimita uma cavidade central, chamada de átrio ou espongiocele que se abre no ósculo. Nas esponjas mais complexas não existe uma cavidade central, mas um labirinto de canais e câmaras revestidas de células flageladas, os coanócitos, chamadas de câmaras vibráteis.
Evolução e relacionamento entre
os tipos de esponjas
Esponjas são um táxon diversificado de animais aquáticos bentônicos de grande importância ecológica, comercial e biofarmacêutica.
Eles são indiscutivelmente o táxon metazoário de ramificação mais antiga e, portanto, eles têm grande significado na reconstrução da evolução inicial dos metazoários como um todo.(Dohrmann et alii, 2012)
No entanto, a filogenia e a sistemática das esponjas ainda estão, em certa medida, não resolvidas, e há um debate em andamento sobre o padrão exato de ramificação de seus clados principais e suas relações com outros animais não bilaterais.
Atualmente reconhece-se para os poríferos cinco classes, uma delas já extinta (Archaeocyata) e quatro ainda existentes (Calcarea Bowerbank, 1864, Demospongiae Sollas, 1885, Homoscleromorpha Bergquist,
1978 e Hexactinellida Schmidt, 1870).
Destas quatro classes a classe Demospongiae é a maior classe contendo 90% de
todas as espécies de esponjas viventes (MURICY et al., 2008). Ao todo fazem parte do filo
cerca de 22 ordens, 128 famílias e 680 gêneros, mas muitos destes táxons mais elevados estão
em discussão devido a novos “insights” obtidos a partir de métodos sistemáticos moleculares
e novas considerações a respeito de suas características morfológicas (MORROW; CÁRDENAS,
2015).
Para reconher-se esses taxa, o estudo das estruturas de sustentação, como fibras de espongina e
espículas, é fundamental na taxonomia de Porifera, uma vez que é com base nestes caracteres
que se determina a classificação destes animais (BERGQUIST, 1978).
In situ pictures of the Mediterranean Homoscleromorpha species from this study. a) Oscarella tuberculata; b) Oscarella lobularis; c) Oscarella viridis; d) Oscarella microlobata; e) Oscarella sp.(researchegate)
Original Caption: Xestospongia muta, the barrel sponge, may live for 100 years and grow to over 6 feet tall. While populations have declined at sites throughout the Caribbean, they appear to be quite healthy on Little Cayman Island. Caribbean Sea, Cayman Islands." Source: NOAA (Public Domain).
Demospongiae morphology and spicule diversity. A. Bath sponge, Spongia officinalis, Greece (photo courtesy E. Voultsiadou); B. Bathyal mud sponge Thenea schmidti; C. Papillae of excavating sponge Cliona celata protruding from limestone substratum (photo M.J. de Kluijver); D. Giant rock sponge, Neophrissospongia, Azores (photo F.M. Porteiro/ImagDOP); E. Giant barrel sponge Xestospongia testudinaria, Lesser Sunda Islands, Indonesia (photo R. Roozendaal); F. Amphimedon queenslandica (photo of holotype in aquarium, photo S. Walker); G. SEM images of a selection of microscleres and megascleres, not to scale, sizes vary between 0.01 and 1 mm. doi:10.1371/journal.pone.0035105.g002.
Diversidade de Hexactinellida
NOVAS ESPONJAS MARINHAS
https://youtu.be/oJeyOU4eSKw
Alternative scenarios for the higher-level relationships of extant sponges. Left: sponge paraphyly (e.g. Sperling et al., 2009). According to this scenario, a sponge-like body plan (white circle) was acquired in the last common ancestor of Porifera and Epitheliozoa (sensu Ax, 1996; Cnidaria, Ctenophora, Placozoa, Bilateria) and subsequently lost (red circle) from the last common ancestor of Epitheliozoa. Alternative paraphyly scenarios exist mainly in earlier studies, where homoscleromorphs were often not included (see text for details). Right: sponge monophyly (e.g. Philippe et al., 2009). According to this scenario, the sponge-like body plan (white circle) was acquired either in the stem lineage of Porifera (P) or, if choanocytes are considered homologous to choanoflagellate cells as judged by outgroup comparison to the well-established sister group of the Metazoa, the Choanoflagellata (see text for details), in stem-group metazoans (S). The latter scenario would require one gain and one loss (indicated by white/red dots marked with S), as in the paraphyly hypothesis (Dohrmann et alii, 2012).
Homoscleromorpha
Homoscleromorpha is a recently recognized class within the phylum Porifera. While molecular analyses support monophyly of Homoscleromorpha, morphological data are largely lacking. Here, we investigate and comparatively analyze the detailed morphology of kinetids (flagellar apparatus) in choanocytes of ten Oscarella species. The kinetid is known to be a phylogenetically important trait that can possibly reflect relationships of sponges at the level of orders. We also conduct a phylogenetic analysis based on previously published sequences. We found the family Oscarellidae is subdivided into two large groups, and choanocyte kinetids structure strongly supports this division. The first group includes Oscarella lobularis, O. tuberculata, O. bergenensis, O. viridis, O. rubra, O. malakhovi, O. microlobata, O. carmela, O. filipoi, and O. zoranja. These species possess nuclei located in a basal position without any connection to the kinetosome. The second group includes O. balibaloi, O. nicolae, O. kamchatkensis, O. pearsei, O. imperialis, and Pseudocorticium jarrei, whose nuclei are connected with the kinetosome. In addition, this second group also shares mesohylar spherulous cells harboring paracrystalline inclusions. The choanocyte kinetid structure seems to be an essential phylogenetic and diagnostic morphological trait, and we propose to include it in future species descriptions. Our results imply assignation of the second group to the genus Pseudocorticium. This taxonomic suggestion, however, needs additional data on the remaining Oscarella species.
Hipótese sobre o relacionamento entre os metazoários e poríferos
A phylogeny of Metazoa adapted from Philippe et al 2009 which considers Porifera monophyletic. Sponge phylogeny is adapted from [48]. Freshwater sponges belong to the group indicated with a box (G4). Letters indicate evidence of epithelial characters: (a) transepithelial resistance (TER) and transepithelial potential (TEP) in freshwater sponges; (b) septate junctions in calcareous sponges; (c) a basement membrane in homoscleromorphs; (d) absence (loss?) of a basement membrane in placozoans; (e) true epithelia with septate junctions, a basal lamina, TER and TEP in cnidarians and bilaterians. Arrows indicate three potential origins of epithelia; the solid bar indicates the most parsimonious scenario for the origin of epithelia (2*)
REPRODUÇÃO
(a) Adult specimen attached to green algae with its basal part, osculum with fringe of spicules at the top. (b) Radial chambers of a juvenile specimen. (c) Syconoid body plan with direction of water flow indicated. (d,e) Fixed and cleared sample containing oocytes. (f) Fixed and cleared sample containing preinversion stage embryos. (g) Cartoon representation of major cell types and embryogenesis stages. (h–j) Histological sections demonstrating key embryogenesis stages. (k) Released live larva (macromeres on top, micromeres to the bottom). (l,m) Olynthus stage juvenile: live specimen and histological section demonstrating major cell types: choanocyte, pinacocyte and porocyte magnified in the inset. Scale bar indicates 5 mm in a, 100 μm in b,d and f, and 10 μm in remaining panels.
Espongiose
Ulisses dos Santos Pinheiro (2008) - Departamento de Zoologia – CCB/UFPE
Espongiose se refere a acidentes com esponjas que normalmente causam inflamações no tegumento atingido. Um dos indícios da espongiose é a coceira, além de forte reação alérgica, a qual pode requerer cuidados médicos urgentes. Isto normalmente ocorre por que as esponjas, ao morrerem, têm seu colágeno completamente decomposto, liberando para o ambiente todo seu esqueleto silicoso que é composto por espículas. Estas espículas penetram no tegumento causando as inflamações. Segundo Machado (1947), os Carajás (uma das tribos indígenas amazônicas) usavam espículas de esponjas de água doce para fabricar cerâmica. Esse autor descreve que “os indígenas pulverizam espongiários, calcinados previamente ou não, e adicionam à argila com que fabricam peças de sua cerâmica”. As mãos das oleiras indígenas que trabalhavam com cauxí (argila mista com espículas de esponjas) possuíam dermatite purulenta. Hilbert (1955) chama a atenção para a preferência dos caboclos pelo caripé (argila mista com cinzas de tecidos vegetais pertencentes a licania, uma rosácea) ao invés do cauxí. Os motivos por essa escolha estavam relacionados com os inchamentos das mãos causadas pelo cauxí e, ainda, porque os utensílios de cozinha fabricados com esse material causavam desarranjos estomacais.
Um dos casos de espongiose mais recente foi o surto de dermatites e cegueiras ocorrido em Araguatins (Tocantins). Sua população ao entrar em contato com a água do rio da região sofriam de vários casos de dermatite e cegueiras. O fato curioso é que estes acidentes só ocorriam na época de águas baixas, não ocorrendo sintomas na época da cheia. Após uma ampla investigação científica, Volkmer-Ribeiro et al. (2006) concluíram que tais lesões eram causadas por duas espécies de esponjas Drulia uruguayensis e Drulia ctenosclera, que ao morrerem liberam na coluna d’água muitas espículas, sendo a gemosclera o principal agente causador de acidentes oculares e dermatites. No caso de Araguantins tal acumulo de espículas foi conseqüência de alterações antrópicas que geraram o desequilíbrio no meio.
Outra espécie de esponjas de água doce que já foram registrados acidentes é a Sterrastrolepis brasilinesis, uma esponjas endêmica da Bacia do Paraná, conhecida popularmente como “espinho de pedra”. Esta espécie possui projeções cônicas que ficam aderidas fortemente ao substrato rochoso e por serem extremamente duras, ao inadvertidamente pisar nestas esponjas, os banhistas acabam perfurando os pés. (ver tb Volkmer-Ribeiro e Batista, 2007)
RESUMINHO
Estrutura Corporal e Sistema Aquífero
• Pinacoderme: superfície externa do corpo da esponja, formada pelos pinacócitos.
• Coanoderme: superfícies internas, compostas por células flageladas, coanócitos; com apenas 1 célula de espessura; pode ser simples e contínua ou dobrada e subdividida formando câmaras;
• Mesoílo: camada intermediária entre pinacoderme e coanoderme; constituído por uma matriz coloidal acelular, com fibras de colágeno, espículas e vários tipos de células; tem papel importante na digestão: transporte de nutrientes, produção de gametas, secreção do esqueleto e excretas por células amebóides.
• Porócitos: poros unicelulares. Neste tipo de célula tubular a agua entra na esponja
• Poros dérmicos ou óstios: pequenas aberturas na pinacoderme;
• Endopinacócitos: células da pinacoderme que recobrem canais internos.
Tipos celulares
1) Células delimitadoras de superfícies:
i. Pinacócitos: formam a pinacoderme – camada contínua superficial externa das esponjas e também revestimento dos canais inalantes e exalantes;
• Endopinacócitos: pinacócitos internos que revestem os canais;
• Exopinacócitos: revestimento externo dos canais;
• Basopinacócitos: células externas da região basal ou de adesão ao substrato – secretam complexo fibrilar de colágeno-polissacarídeo = lâmina basal;
• Em esponjas dulcícolas, basopinacócitos são ativos na nutrição, lançam filapódios, semelhantes às amebas.
ii. Porócitos: células cilíndricas, como tubos, localizadas na pinacoderme – forma os óstios (Lat. Ostium = porta, entrada);
• São contráteis – podem abrir e fechar o poro – regulação do diâmetro dos óstios;
iii. Coanócitos:
célula flageladas com colarinho, formam a coanoderme e criam correntes hídricas no sistema aquífero.
• Longo flagelo é rodeado pelo colar com 20 a 55 microvilos citoplasmáticos;
• Coanócitos localizam-se sobre o mesoílo (internamente ou em câmaras);
• Atuam na captura de partículas alimentares (fagocitose e pinocitose);
Habitat
Marinho e dulcícola
Nutrição
Heterotrófico por filtração
Fototrófica (simbiose com bactérias)
https://youtu.be/Y4uc9cSolDk
PORIFEROS E CNIDARIOS
Exercícios
1) (10) Faça um esquema do corpo de uma esponja em seu caderno e coloque os nomes nas estruturas constituintes e dê a função de cada uma dessas estruturas.
2) (2) Descreva a digestão nesses organismos. (Faça um esquema em seu caderno da digestão intracelular nesses organismos e explique todo o processo).
3) (2) Cite a apomorfia desses animais e faça um esquema da mesma e dê os nomes das estruturas.
4) (4) Que características ancestrais os organismos do subreino PARAZOA compartilham com os EUMETAZOA?
5) (5) Construa um cladograma dos Metazoários e coloque as apomorfias relativo a cada ramo (clado) existente.
6)(4) Leia o texto abaixo: "Espongiose" de Ulisses dos Santos Pinheiro (2008), extraído de um importante trabalho sobre esponjas publicado na Revista Eletrônica do Museu Nacional e USP Ciências e responda as seguintes questões:
a) O que tem nas esponjas que pode causar irritações nos tecidos humanos?
b) Após ler o texto, pesquise para que serve a sílica atualmente para nós humanos.
c) Como é chamada a doença causada pelas esponjas
7) (4) O tamanho e a forma das esponjas variam muito, entretanto, o fluxo de água no interior das espojas sempre segue o seguinte sentido: poros → átrio → ósculo.
Baseando-se nas informações acima, no esquema e no seu conhecimento sobre o modo de vida dos poríferos, explique a importância ecológica das esponjas para a saúde dos ambientes aquáticos, indicando o tipo mais eficiente, justificando a indicação.
8) (3) Uma esponja é capaz de movimentar um volume de água considerável. Estudos mostram que algumas esponjas grandes (2m3 de volume e um poro com 50 micrômetros de diâmetro) podem bombear seu próprio volume em água a cada 20 segundos. Sendo assim, que volume de água essa esponja bombeará em 1minuto?
9) (5) Descreva através de um esquema o desenvolvimento embrionário dos metazoários, salientando sua apomorfia.
Bibliografia
BARNES, R. Invertebrate Zoology (modificado)
CAMPBELL; N. RICE, J.B. e cols. 2010. Biologia. Porto Alegre, Artmed Ed. S.A. ERESKOVSKY, Alexander V. The Comparative Embryology of Sponges Dordrecht; St. Pertersburg Univ. Press; Springer Science, 2010.
(1*) The evolution of animals (metazoans) from their unicellular ancestors required the emergence of novel mechanisms for cell adhesion and cell–cell communication. One of the most important cell adhesion mechanisms for metazoan development is integrin-mediated adhesion and signaling. The integrin adhesion complex mediates critical interactions between cells and the extracellular matrix, modulating several aspects of cell physiology. To date this machinery has been considered strictly metazoan specific. Here we report the results of a comparative genomic analysis of the integrin adhesion machinery, using genomic data from several unicellular relatives of Metazoa and Fungi. Unexpectedly, we found that core components of the integrin adhesion complex are encoded in the genome of the apusozoan protist Amastigomonas sp., and therefore their origins predate the divergence of Opisthokonta, the clade that includes metazoans and fungi. Furthermore, our analyses suggest that key components of this apparatus have been lost independently in fungi and choanoflagellates. Our data highlight the fact that many of the key genes that had formerly been cited as crucial for metazoan origins have a much earlier origin. This underscores the importance of gene cooption in the unicellular-to-multicellular transition that led to the emergence of the Metazoa.(Pedrós et alii, 2010)
(2*) Epithelial tissue — the sealed and polarized layer of cells that regulates transport of ions and solutes between the environment and the internal milieu — is a defining characteristic of the Eumetazoa. Sponges, the most ancient metazoan phylum [1], [2], are generally believed to lack true epithelia [3], [4], [5], but their ability to occlude passage of ions has never been tested. Here we show that freshwater sponges (Demospongiae, Haplosclerida) have functional epithelia with high transepithelial electrical resistance (TER), a transepithelial potential (TEP), and low permeability to small-molecule diffusion. Curiously, the Amphimedon queenslandica sponge genome lacks the classical occluding genes [5] considered necessary to regulate sealing and control of ion transport. The fact that freshwater sponge epithelia can seal suggests that either occluding molecules have been lost in some sponge lineages, or demosponges use novel molecular complexes for epithelial occlusion; if the latter, it raises the possibility that mechanisms for occlusion used by sponges may exist in other metazoa. Importantly, our results imply that functional epithelia evolved either several times, or once, in the ancestor of the Metazoa.
Structure of the sponge epithelium.
(A) Diagram of the experimental chamber, and (B) scanning electron microscopy (SEM) of sponge tissue showing: the bilayered dermal tissues (dt), suspended over the subdermal space (ss) by shafts of spicules (s); the choanosome (ch) with the choanocyte pumps; excurrent canals (ec); and an osculum (o). For transepithelial recordings, apical (AP) and basolateral (BL) compartments were effectively separated by two confluent cell layers - the basopinacoderm (bp) and endopinacoderm (enp) - surrounding a thin mesohyl (me) (inset); scale bar b, 100 µm. (C) SEM of the dermal tissue shows exopinacocytes are very close to one-another (opposing arrows show cell-cell spacing); scale 1 µm. (D) Fluorescent labelling of the basopinacoderm with the steryl dye FM 1–43 highlights the borders of cells showing that a confluent layer of cells covered culture membranes; scale 10 µm. (E) Transmission electron microscopy of exopinacocytes shows tight membrane apposition between adjacent cells (arrow). These areas were associated with 10 nm diameter cytoskeletal fibres (arrowheads); scale 200 nm. (F) Fluorescent labelling using phallacidin reveals dense plaques of actin (arrowhead) between cells; nuclei, blue; scale, 5 µm.































{kind=link}



{kind=link}












{kind=link}

2 comentários:
Muito boa matéria, parabéns!
Muito boa á matéria! Parabéns"
Postar um comentário