As proteínas são algumas das macromoléculas biológicas mais abundantes e diversas que ocorrem em todas as células e em todas as partes das células. Elas possuem uma enorme diversidade de funções biológicas. São os instrumentos moleculares por meio dos quais a informação genética é expressa.
As subunidades básicas das proteínas são os aminoácidos. São 20 os aminoácidos primários, que constituem uma espécie de alfabeto para virtualmente todas as proteínas encontradas na natureza.
Todos os aminoácidos comuns são α-aminoácidos porque possuem um grupo carboxila e um grupo amino ligados ao mesmo átomo de carbono (o carbono α). Eles diferem entre si nas suas cadeias laterais, ou grupos R, que variam em estrutura, tamanho e carga elétrica e que influenciam a solubilidade dos aminoácidos em água.
De acordo com as propriedades dessa cadeia lateral, os aminoácidos podem ser divididos em: grupos R alifáticos não-polares; grupos R aromáticos; grupos R não-carregados polares; grupos R carregados positivamente; grupos R carregados negativamente. Exceto na glicina, os carbonos α de qualquer aminoácido são centros quirais, o que implica na existência de pelo menos dois estereoisômeros para cada aminoácido. Quando processados por organismos, normalmente apenas um dos isômeros é encontrado, pois as enzimas envolvidas na síntese dos aminoácidos são estereoespecíficas. São os L-aminoácidos (chamados assim por possuírem conformação análoga a do L-gliceraldeído). (Nelson et al, 2011).
Macromoléculas
As proteínas são macromoléculas formadas por um ou mais (n) peptídeos ou aminoácidos, unidos por uma ligação peptídica formando um polímeros de aminoácidos, os quais estão arranjados de maneira particular para cada proteína. Tanto a ordem dos aminoácidos (na cadeia polipeptídica) quanto a estrutura depende da proteína e da função que ela exerce e é determinada geneticamente pelo DNA.
As proteínas são sintetizadas como uma seqüência linear de aminoácidos unidos em uma estrutura poliamida (polipeptídio) mas assumem uma configuração tridimensional complexa para que possam realizar suas funções. Algumas proteínas só conseguirão alcançar sua forma funcional ao ligarem-se a cofatores como uma vitamina. (DNA-mRNA-Traducao - proteína enzimática ou apoenzima + cofator (vitamina) = Holoenzima.
Essa estrutura foi esclarecida por Emil Fischer (1852-1919), que verificou serem esses compostos formados pela condensação entre muitas moléculas de (α) alfa-aminoácidos através de ligações denominadas ligações peptídicas. São chamamos de alfa-aminácidos por que possuem um carbono alfa (como dito acima).
Carbono alpha (alfa)
Características básicas de uma ligação peptídica (entre dois aminoácidos), sucintamente, são:
Origina cadeia (poli)peptídica formada por resíduos de aminoácidos.
Origina duas extremidades, uma N-terminal (aminoterminal) e outra C-terminal (carboxila terminal).
Para que acontecça uma ligação peptídica ou condensação é necessário uma complexa maquinaria celular, como os ribossomos, mRNA, tRNA, enzimas, proteínas e energia.
A hidrólise é feita por proteases, ácidos e bases fortes, t°↑;
Ressonância: apesar de ser representada por 1 único traço, a ligação peptídica, possui características intermediárias entre uma ligação simples e uma ligação dupla. Átomos que participam da ligação peptídica (H-N-C-O) formam 1 plano rígido. A rotação só é possível em torno do carbono alpha (Cα1). (modif. Infoescola, preparaenem).
As proteínas apresentam peso molecular extremamente elevado, e são compostas por carbono, hidrogênio, nitrogênio e oxigênio, sendo que praticamente todas elas possuem enxofre. Elementos como ferro, zinco e cobre também podem estar presentes.
Todas as proteínas são formadas por um conjunto de 20 aminoácidos, arranjados em sequências específicas determinadas pela ordem dos nucleotídeos no DNA (gene).
Proteínas presentes no envelope viral do SARS-CoV-2
Aminoácidos
Os aminoácidos são moléculas orgânicas que apresentam um carbono saturado, denominado de carbono alfa (central), que realiza uma ligação com um átomo de hidrogênio, uma ligação com um grupamento amino (NH2), uma ligação com um grupamento ácido (COOH), e uma ligação com um radical orgânico qualquer, R, também chamada de cadeia lateral. Esta esta última ligação é diferente para cada um dos vinte (20) aminoácidos e é a que distingue um aminoácido de outro.
Funções de alguns aminoácidos
Leucina, isoleucina e valina
São aminoácidos conhecidos como BCAA, estão envolvidos na reparação muscular, aumento de proteínas e durante atividades físicas ajudam na produção de energia.
Arginina
Um aminoácido ligado a melhora da memória, ajuda na resistência física, aumentando o desempenho durante a atividade física. É utilizada como suplemento para treinos musculares.
Alanina
Aminoácido também envolvido no metabolismo para obtenção de energia.
Treonina
Aminoácido importante na síntese de colágeno e elastina que são proteínas estruturais.
Metionina
Aminoácido envolvido na resposta imunológica do nosso corpo, sua falta pode ocasionar a queda de cabelo.
Triptofano
Aminoácido envolvido na síntese ou produção de outros aminoácidos, alguns pesquisadores afirmam que bons níveis de serotonina associado com bons níveis do triptofano garantem um nível estável de ânimo em uma pessoa, contribuindo contra a depressão.
Há também alguns aminoácidos especiais, que aparecem em algumas proteínas específicas. É o exemplo da N-metilarginina e N-acetillisina encontradas nas histonas.
Aminoácidos essenciais e não essenciais
Ainda que nosso organismo seja composto por cerca de 250 mil proteínas diferentes, cada uma com funções específicas em momentos e células específicas, todas estas proteínas são formadas por apenas 20 aminoácidos.
Destes vinte aminoácidos, nosso corpo é capaz de fabricar somente onze (11) destes 20. Os outros nove, que são a histidina, isoleucina, leucina, lisina, metionina, fenilalanina, treonina, triptofano e valina, são os chamados aminoácidos essenciais.
Estes aminoácidos não podem ser sintetizados pelas nossas células (humanos e os animais monogástricos não sintetizam os aa essenciais), assim, temos que obtê-los por meio da alimentação.
Daí a importância de uma alimentação adequada que disponibilize todos os 20 aminoácidos que nosso corpo necessita. É importante obtermos esses aminoácidos de forma simultânea, e aqui encontramos uma dificuldade, porque a maioria dos alimentos não contém individualmente todos os aminoácidos essenciais em quantidade suficiente.
Alguns alimentos como as carnes magras, os ovos, o leite e seus derivados (queijo, iogurte, coalhada) contêm os nove aminoácidos essenciais e também os onze (11) não essenciais que nosso organismo requer para funcionar corretamente. Os alimentos de origem animal têm proteínas de alto valor biológico e nos fornecem todos os aminoácidos essenciais em conjunto.
Alimentos como carne de porco, frango e bovina, além de peixes como salmão, mero, atum e sardinha são ricos em isoleucina, valina, leucina, fenilalanina, treonina, metionina, histidina e lisina.
Frango, peru, coelho e peixes como salmão, sardinha, vieira, bacalhau e atum são ricos em triptofano.
O leite e seus derivados, em especial o queijo, também contêm praticamente todos os aminoácidos. E o ovo, especialmente, a clara, contém aminoácidos como isoleucina e valina.
Apesar de alimentos de origem animal apresentarem todos os aminoácidos (essenciais e não essenciais) simultaneamente, junto com a carne estamos ingerindo uma porção extra de gorduras, que nem sempre é bom para o nosso organismo. Assim, devemos e podemos lançar mão dos alimentos de origem vegetal como o grão-de-bico, a soja, alguns feijões, trigo-sarraceno, quinoa, amaranto, sementes de cânhamo e pistache contêm todos os aminoácidos essenciais. Todavia, lentilhas têm uma quantidade limitada do aminoácido essencial metionina, e os cereais, como o arroz integral, por exemplo, contêm pouca lisina e treonina. Portanto, se fizermos um prato que combine lentilhas (ricas em lisina, pobres em metionina) com arroz integral (rico em metionina e pobre em lisina), obteremos uma refeição com proteínas completas. (De La Nuez, 2019).
Não há necessidade de combinar esses alimentos na mesma refeição. Isso quer dizer que você pode comer lentilhas no almoço e arroz integral no jantar, já que o fígado vai estocar e oferecer ao nosso corpo os aminoácidos essenciais quando for necessário. (Veja essa reportagem: Cogumelos e memória para discutirmos)
Os aminoácidos são imprescindíveis para a vida. Ao ingerirmos proteínas de origem animal e vegetal estamos suprindo nosso corpo com os tijolos, os aminoácidos que servirão para construção de novas células, para reparo de danos, para defesa, para transporte e coordenação entre outras funções.
Um alimento que apresentar todos os 20 aminoácidos necessários para formar as proteínas existentes e imprescindíveis para o com funcionamento do nosso organismo, é chamado de alimento completo. Alimentos de origem animal, como carne, leite, queijo, peixe e ovos, possuem proteínas de alta qualidade, com todos os aminoácidos necessários para o nosso organismo.
Aminoácidos essenciais
Aminoácidos essenciais
Cada um dos aminoácidos essenciais tem funções específicas e vitais para o bom funcionamento dos organismos. Eles são necessários para a construção de tecidos, músculos, alguns hormônios e enzimas. Devem necessariamente ser obtidos por meio da alimentação ou da suplementação. (Essentialnutrition,2020).
Um aminoácido essencial, ou aminoácido indispensável, é um aminoácido que não pode ser sintetizado "de novo" (do zero) pelo organismo com rapidez suficiente para suprir sua demanda e, portanto, deve ser obtido da dieta. Dos 21 aminoácidos comuns a todas as formas de vida, os nove aminoácidos que os humanos não podem sintetizar são fenilalanina, valina, treonina, triptofano, metionina, leucina, isoleucina, lisina e histidina.
Fenilalanina: este aminoácido potencializa a produção de hormônios e neurotransmissores, como a dopamina, adrenalina e noradrenalina, substâncias ativadoras do sistema nervoso central e periférico. Ele é importante para manter atividades mentais e psicológicas em equilíbrio e melhora a capacidade cognitiva.
Valina: é um aminoácido que auxilia na síntese de outros aminoácidos, no reparo dos tecidos e na manutenção do balanço nitrogenado equilibrado.
Leucina: estimula as sínteses das proteínas musculares e é um dos principais combustíveis das reações anabolizantes.
Treonina: responsável pela produção de mucina, proteína necessária para a manutenção da integridade e função intestinal. Importante para a imunidade e responsável pela síntese de glicina.
Triptofano: substância que incentiva a liberação de serotonina (hormônio que regula funções importantes, como o humor, a temperatura do corpo e ritmo cardíaco) e a melatonina (hormônio relacionado ao sono).
Metionina: é uma importante fonte de enxofre e outros compostos necessários para o metabolismo e componentes celulares. A metionina se converte em cistina, aminoácido importante na síntese de glutationa (antioxidante natural).
Isoleucina: responsável pela síntese de glutamina e alanina, além de agir no desenvolvimento e reparos dos músculos.
Lisina: trata-se de um aminoácido que regula a produção de óxido nítrico, é componente estrutural do colágeno e possui ação antiviral.
Histidina: A histidina é um aminoácido essencial provavelmente pela quantidade muito pequena (insuficiente) que nosso corpo pode produzir. Apresenta uma cadeia lateral composta por um anel imidazol carregado positivamente a níveis de pH fisiológico. A histidina é essencial para as crianças, já para os adultos podem sintetizá-la, embora não esteja totalmente claro se a quantidade que é produzida é suficiente para cobrir todas as necessidades e por isso é considerado essencial.
Esta ligado ao crescimento e regeneração de tecido entre eles o tecido muscular manutenção das células cerebrais
É encontrado na natureza como o enantiômero levorrotatório da histidina, i.e., a L-histidina. (Breve explicação sobre enantiomeria mais abaixo em Curiosidades da vida).
A L-histidina é um aminoácido presente em diversos alimentos, tanto de origem animal como vegetal, como ovos, leite, carne bovina, suína, peixe (robalo), soja, feijão, quinoa, ervilha. A L-histidina é o aminoácido precursor na síntese de outros compostos biológicos como a histamina que atua com um modulador imunológico (esta ligado a reações como urticaria, edema, bronco constrição, secreção mucosa, aumento da permeabilidade vascular, lacrimejo, tosse, ou seja, reações alérgicas, ou resposta imunológica do nosso corpo), e em combinação com β-alanina a histidina se converte em carnosina um potente antioxidante endógeno (um dipeptídeo encontrado no tecido esquelético, muscular, tecido nervoso (cérebro) e coração). A carnosina auxilia na diminuição do nível de acidez dentro das células musculares (essa acidez é causada pela liberação de ácido lático durante o exercício muito intenso), retardando a fadiga.
A carnosina, também é necessária para a atuação de uma enzima chamada óxido nítrico sintetase, responsável pela produção do óxido nítrico (NO2). Uma vez na circulação, o óxido nítrico promove a dilatação dos vasos sanguíneos, o que aumenta o fluxo sanguíneo para dentro do tecido muscular. Isso significa mais oxigênio, aminoácidos, açúcares e nutrientes chegando nas células musculares, permitindo uma contração prolongada e uma aceleração na recuperação pós-treino.
A histidina está envolvida na catálise de diversas reações bioquímicas em nossas células, como a metilação dos resíduos de arginina, síntese de ácido urocânico, é o precursor da síntese de ácido glutâmico, protetor contra a radiação UV (ultravioleta) na pele e imuno-regulador, além da síntese de compostos orgânicos intermediários genômicos e não genômicos. (hsnstore)
O ácido urocânico: por toda a extensão da nossa pele existe um manto ou filme hidrolipídico com função de protetor solar nativo da pele. O manto hidrolipídico é um biofilme protetor contra radiações UV. É uma barreira protetora que mantém toda extensão da camada mais externa da pele naturalmente hidratada. É também conhecido como barreira cutânea, e é como uma película finíssima, um biofilme formado por uma emulsão de compostos hidrofílicos (amigos de água) e lipofílicos (amigos de gordura). Esta emulsão contém diversas substâncias importantes para a defesa e saúde da pele, como o ácido urocânico. O ácido urocânico é uma tipo de protetor solar produzido pelo nosso organismo. Quando tomamos sol, antes mesmo do nosso corpo começar a fabricar a melanina, que também é um tipo de protetor, que protege a pele contra a radiação solar direta, nossas glândulas sudoríparas já estão em pleno funcionamento, produzindo o ácido urocânico.
Enquanto o filme hidrofílico atrai e absorve a água para as camadas superficiais da pele, o composto lipofílico atrai e segura gordura. Juntos, eles criam um biofilme semioclusivo que impede a saída excessiva de água, ajudando a manter a pele hidratada. Além disso, uma tarefa importante que ele desempenha é conservar o pH da pele nos níveis adequados. Essa emulsão hidrolipídica possui toda uma família de substâncias que garante o ambiente ácido da pele, o que é fundamental para inibir a permanência e a proliferação de bactérias e fungos que tentam se estabelecer nessa camada superficial da pele e eventualmente invadir nosso corpo. (riobiocosméticos, mundoboaforma).
A histidina e a Hemoglobina A (HbA)
A hemoglobina que predomina nos eritrócitos de um ser humano adulto (HbA), possui duas cadeias polipeptídicas (um par idêntico) que contêm 141 aminoácidos e são denominadas cadeias alfa (α). O par restante de cadeias idênticas contém 146 aminoácidos, e são denominadas cadeias beta (β). Essas cadeias formam junto um tetrâmero.
A hemoglobina possui quatro grupos heme, cada heme é formada por um anel protoporfirínico contendo um átomo de ferro. A protoporfirina III diferencia-se das demais pelas cadeias laterais presentes nos anéis pirrólicos.
As ligações covalentes entre a heme e a globina ocorrem entre as histidinas da globina e o radical propil do pirrol. O ferro liga-se a globina através da histidina proximal e é responsável por carregar uma molécula de oxigênio através de uma ligação fraca com histidina distal (oxihemoglobina).
A ligação com gás carbônico se faz através de grupamentos aminas laterais da globina originando a carbohemoglobina, embora a maior parte do CO2 seja eliminada dos tecidos na forma de íon bicarbonato.
O monóxido de carbono (CO) tem alta afinidade pela hemoglobina e sua ligação origina a carboxihemoglobina que é irreversível. (PA).
AMINOÁCIDOS ESSENCIAIS E NÃO-ESSENCIAIS OU NATURAIS
Curiosidades do fenômeno Vida
1) Quiralidade
Ao sintetizarmos moléculas de aminoácidos e acúcares em
laboratório, se faz um conjunto, uma mistura das duas nas mesmas proporções:
metade L, de levógeras, que são as moléculas "canhotas", e metade D, de dextrógeras, as moléculas "destras".
Ao abordarmos aspectos da quiralidade, torna-se importante situarmos o termo num contexto mais amplo de “isomeria”. Isômeros são compostos que possuem os mesmos constituintes atômicos, porém suas disposições na molécula são diferentes, conferindo conseqüentemente características químicas diversas.
Os isômeros possuem diferentes classificações de acordo com esta distribuição atômica (isômeros de função, isômeros de cadeia, etc) e, o que nos interessa aqui, é a distribuição de seus átomos no espaço: os estereoisômeros.
Estereoisômeros são aqueles isômeros cujos átomos ou grupos de átomos possuem uma distribuição espacial diferente na molécula. Eles podem ser divididos em geométricos ou ópticos.
Os isômeros geométricos são estereoisômeros que não apresentam atividade óptica* e sua terminologia está centrada em cis (do mesmo lado) e trans (lados opostos) para descrever sua disposição espacial. Para os alquenos, na maioria da vezes, pode-se igualmente falar em Z e E para cis e trans, respectivamente. Isômeros ópticos são aqueles que apresentam atividade óptica, possuindo centros quirais ou centros assimétricos. Dentro deste conceito, podemos dizer que a quiralidade manifesta-se quando a molécula possuir uma das seguintes características:
a) um centro de quiralidade* (C*, Si*, S*, P*, N*);
b)
um eixo de quiralidade (alenos, espiranos);
c) um plano de
quiralidade e
d) uma forma de hélice (hexaeliceno).
As moléculas com um elemento de quiralidade apresentam
enantiomeria. Os enantiômeros são estereoisômeros relacionados entre si por uma simetria em relação a um plano e são
também conhecidos como antípodas ópticos .
As moléculas que apresentam dois ou mais elementos de
quiralidade apresentam a diastereoisomeria. Diastereoisômeros
não são enantiômeros; por exemplo, para uma molécula com
dois centros quirais, teremos neste caso dois diastereoisômeros
e todos os seus quatro isômeros não serão sobreponíveis no
espelho plano4,5 Enquanto que os enantiômeros possuem as
mesmas características físicas, como solubilidade ou ponto de
fusão, os diastereoisômeros podem apresentar diferentes propriedades físicas e também químicas, aspectos que veremos
mais adiante.
Em todas substancias e tudo o
que existe na natureza, as formas L e D deveriam
sempre coexistir, se misturar, formando uma mistura racêmica.
O surpreendente, quando olhamos o
organismo humano, é que todos os
aminoácidos são L, não temos nenhum
D. Daí analisamos os açúcares, que também deveriam ter L e D, mas todos os açúcares são D
e nenhum L.
Como explicar isso num
mundo todo assimétrico, aquiral, onde
sempre deveríamos encontrar uma mistura dos dois?”, questiona Eberlin. Não
existe (ou não existia) nenhuma explicação lógica, dentro da ciência, para que
se privilegiasse uma dessas formas. Como
explicar essa separação do D para os
aminoácidos e do L para os açúcares na
formação de seres vivos?
Espectrometria de massas. A espectrometria de massas é uma técnica de análise instrumental da química em que se
visualiza com precisão o universo molecular. Foi por meio dela que as equipes
de Marcos Eberlin e de Robert Cooks realizaram experimentos, detectando algo
inédito: “Pegamos uma mistura L e D de
um aminoácido e conseguimos colocar no
L uma marca química, distinguindo-o do
D. Depois, marcamos dois.
Percebemos
então que os L e D se agrupavam naturalmente: os D de um lado, formando uma
estrutura cilíndrica, e os L para outro, formando outra estrutura cilíndrica. Foi bastante interessante, pois nunca se pensou
que esse processo de separação pudesse
ocorrer naturalmente”, lembra Eberlin.
Era um arranjo geométrico tridimensional especial. Como ilustração, o pesquisador da Unicamp recorre às brincadeiras de roda: “Se alguém for brincar
virado de costas ou dando as mãos invertidas, não vai se encaixar na roda. O
mesmo se dá com os aminoácidos, que
se agrupam porque a estrutura é como a
de uma roda: somente aqueles que dão
a mão corretamente se unem, somente
os L (Levógeros, virados para a esquerda) ou os D
(dextrógeros, virados para a direita)”.
O projeto na Unicamp sobre técnicas
modernas em espectrometria de massas e
suas aplicações em química e bioquímica
conta com financiamento da Fapesp e, na
Universidade de Purdue, da agência
National Science Fundation (NSF). Eberlin
ressalta que talvez se tenha proposto uma
explicação apenas para o primeiro passo
do processo de homoquiralidade dos seres vivos, de como separar naturalmente
os aminoácidos. A segunda etapa, da seleção, possivelmente nunca será explicada
na totalidade.
Na mesma pesquisa, comprovou-se também a propagação desse processo de separação para outros aminoácidos, visto que o organismo não possui
apenas um, mas vinte aminoácidos. “Quando nada se tem, o primeiro passo é extremamente importante”, justifica.
Quiralidade e enantiomeria
É encontrado na natureza como o enantiômero levorrotatório da histidina, i.e., a L-histidina. (Breve explicação sobre enantiomeria)
(S) - (+) - ácido láctico (esquerda) e (R) - (-) - ácido láctico (direita)
são imagens espelhadas (especulares) não sobreponíveis um do outro.
Cada composto S e R desse par são chamados enântiomorfos (IUPAC ref. 6).
Um enantiômero existe sob forma de dois pares de isômeros, não superponíveis mas simétricos em relação a um plano (figura acima).
No caso do ácido lático (ácido-2-hidróxi-propanoico) observe que seus quatro ligantes são diferentes. Além disso, por ter em sua estrutura um carbono assimétrico (quiral), ele possui dois isômeros que são a imagem especular um do outro, sendo chamados de enantiômeros. A figura a seguir mostra que esses dois enantiômeros guardam entre si uma relação objeto-imagem especular de modo que não podem ser sobrepostas.
Os dois enantiômeros do ácido lático são opticamente ativos, sendo então chamados também de enântimorfos (do grego enantios, que significa oposto; e morphos, que é forma; isto é, formas opostas) ou antípodas ópticos, pois ambos desviam o plano de luz polarizada com um mesmo ângulo, mas em sentidos opostos, o que torna a mistura de ambos opticamente inativa.
Possuir um carbono quiral resulta em um deles desviar o plano de luz polarizada para a direita, sendo chamado de isômero dextrógiro (d) (do latim dexter, direito); enquanto que o outro desvia para a esquerda, sendo designado de isômero levógiro (l) (do latim laevus, esquerdo). Um é conhecido como ácido L (l), (+) láctico ou ácido (S) lático; e o outro que é sua imagem refletida no espelho, é o ácido D (d), (-) láctico ou ácido lático (R). Uma mistura dos dois em quantidades iguais é chamada de ácido DL-láctico ou ácido láctico racêmico.
O ácido láctico é higroscópico (capaz de absorver umidade do ar).
A tabela a seguir mostra que os dois ácidos láticos têm todas as propriedades físicas e químicas iguais, exceto as propriedades fisiológicas por causa da diferença no desvio do plano da luz polarizada (preparaenem).
O ácido DL-láctico é miscível com água e com etanol acima do seu ponto de fusão, que é em torno de 16 ou 18 °C. O ácido D-láctico e o ácido L-láctico (separados) têm um ponto de fusão mais elevado (52,8 °C). O ácido láctico produzido pela fermentação do leite é freqüentemente racêmico. Algumas espécies de bactérias produzem apenas ácido (R) lático. Por outro lado, o ácido láctico produzido pela respiração anaeróbia nos músculos dos animais tem a configuração (S)ou (L) e às vezes é chamado de ácido "sarcolático", do grego: sarx, sarcodes, σάρξ, sarx que significa carne.
Enantiômeros
Enantiômeros do grego: ἐνάντιος, enántios: oposto; e μέρος, méros: parte; tambem chamado de isômero optico, antipoda, ou antípoda óptica. Enantiômeros são moléculas especulares uma da outra. i.e., são imagens vista no espelho uma da outra. Não são sobreponíveis, nem por rotação, nem por translação. A mistura de enantiômeros numa solução denomina-se mistura racêmica. Enantiômero é um de dois estereoisômeros de um composto quiral. A maioria das moléculas presentes na estrutura dos organismos vivos são quirais. Por exemplo, dos vinte aminoácidos do nosso organismo, 19 estão na forma L e o único que não está na forma L é a glicina, só porque não apresenta um carbono quiral. Desta forma percebemos a importância da quiralidade em organismos vivos. Os estereoisômeros que têm as mesmas propriedades físicas e químicas, exceto pela interação com o plano de luz polarizada ou com outras moléculas quirais. Desta forma possuem atividade biológica diferente, mesmo possuindo ligações químicas e distâncias semelhantes. Usando aluz polarizada. Se após atravessar o composto orgânico, o plano da luz polarizada desviar-se no sentido horário, para a direita, existe ali o isômero dextrogiro. Entretanto, se a luz polarizada for desviada, no sentido anti-horário, i.e., para a esquerda, há o isômero levogiro. Ambos, portanto, são opticamente ativos e são chamados de enantiômeros ou antípodas ópticos ou enântimorfos, pois desviam a luz polarizada de um mesmo ângulo, mas em sentidos opostos. Assim, se na mistura existir ambas as formas em igual quantidade, quando a luz polarizada atravessar esse composto, um anulará a atividade óptica do outro. A essa mistura damos no nome de mistura racêmica, que é opticamente inativa. (W1). Atividade óptica: é a capacidade que certas substâncias possuem de desviar o plano da luz polarizada. Se o desvio for para a direita ela será dextrorotatória (latim: dextro, direita) adotando o sinal (+) ou D ou d, se for para a esquerda, levorotatória (latim: laevus = esquerda) e adota o sinal (-), L ou l. (Lima, 1997). Segundo esta mesma pesquisadora os enantiômeros apresentam desvios polarimétricos opostos de mesma magnitude:
+ α (+) ou d (dextro); - α (-) ou l (levo)
onde o ângulo de rotação α é simplesmente a rotação observada. Misturas contendo igual quantidades de enantiômeros (50:50) são chamadas de misturas racêmicas (ou racematos) e são opticamente inativas. A falta de atividade óptica, neste caso, é decorrente do fato de que enquanto um dos enantiômeros desvia o plano da luz para um determinado valor, o seu par o desvia, na mesma proporção, na direção exatamente oposta, anulando o resultado final. Este é o caso em que uma substância quiral é inativa opticamente (Lima, 1997).
O químico sueco Carl Wilhelm Scheele foi a primeira pessoa a isolar o ácido láctico em 1780 do leite azedo. O nome reflete a forma de combinação de lact, derivada da palavra latina lac, que significa leite. Em 1808, Jöns Jacob Berzelius descobriu que o ácido láctico (na verdade, L-lactato) também é produzido nos músculos durante o esforço. Sua estrutura foi estabelecida por Johannes Wislicenus em 1873. Em 1856, o papel do Lactobacillus na síntese do ácido láctico foi descoberto por Louis Pasteur. Essa via foi usada comercialmente pela indústria farmacêutica alemã Boehringer Ingelheim em 1895 (W2).
O nome “mistura racêmica” veio do latim racemus, que significa cacho de uva. Isso porque o cientista Louis Pasteur descobriu cristais de ácido tartárico produzidos na fermentação do suco de uva na produção do vinho. Ao estudá-los, ele viu que alguns desviavam o plano de luz para a direita, mas outros eram inativos, não provocando o desvio da luz polarizada. Isso acontecia porque, na verdade, esse segundo não era um isômero do ácido tartárico, mas, na verdade, uma mistura racêmica. Ele viu também que era possível a partir dessa mistura obter o isômero levogiro com o uso do lêvedo, pois ele só consome o isômero dextrogiro, deixando o levogiro. (prepraenem).
Patrick (1995), em seu livro "An Introduction to Medicinal Chemistry", resume muito bem as duas importantes características dos enantiômeros: “Existem somente duas diferenças detectáveis entre dois enantiômeros de um composto quiral: eles desviam o plano da luz polarizada em direções opostas e ainda interagem diferentemente com outros sistemas quirais, como as enzimas. Isso tem sido de grande importância para a indústria farmacêutica.” (Patrick, G. L.. An Introduction to Medicinal Chemistry. Oxford, Oxford University Press, 1995).(Lima, 1997).
Teorias obscuras. Na opinião de Eberlin(2002) todos que tentam explicar a homoquirogênese (a criação da homoquiralidade
dos seres vivos) de certa forma usam teorias um tanto “obscuras”, processos físicos
como ação de luz polarizada, campo magnético da terra e separação na superfície de
cristais. “São teorias difíceis de provar ou
contestar. São fundamentos meio esotéricos, como a de que os aminoácidos
quirais teriam surgido em outro planeta e
trazidos para a Terra por um
cometa. É uma questão mais
de fé do que de razão, em que
se acreditava ou não. Este é o
primeiro mecanismo químico
relacionado com a homoquiralidade e, consequentemente, com as teorias sobre
a origem da vida”.
Os pesquisadores e cientistas, ao procurarem entender a arquitetura
química dos seres vivos, adquirem maior conhecimento
de nosso corpo, ampliando as
condições de melhor cuidar
dele, preconiza Marcos Eberlin. (modif. UEL,2002).
Para vc que chegou até aqui, um mistério para resolver
2) Lewis Carroll, em sua segunda obra-prima Alice no País dos Espelhos (1872), narra, no primeiro capítulo, o diálogo da personagem Alice com a sua gatinha Mimi: " – E, se não se corrigir já e já, eu a atiro para dentro da Casa do Espelho. [...] é uma sala igual à nossa, só que as coisas estão todas invertidas [...] . E lá lhe dariam seu leite? Quem sabe se o leite do Espelho nãoé bom para beber ..."CARROLL, L. Alice no País dos Espelhos. São Paulo: Martin Claret, 2007. p. 19.
a) As proteínas do leite da Casa do Espelho poderiam servir de alimento para Mimi? Justifique a sua resposta.
b) Sabendo que a lactose [4-O-(β-D-galactopiranosil)-β-D-glicopiranose] é um dissacarídeo, estabeleça uma metodologia para a separação da lactose do mundo real e da Casa do Espelho por HPLC, incluindo a escolha da coluna, da fase móvel e do detector.
A palavra anfótero vem do grego amphoteroi (ἀμφότεροι) que significa: ambos, ou um e outro. Uma substância de caráter anfótero é aquela capaz de reagir com comportamento ácido ou básico. Substâncias anfotéricas podem ser moléculas, íons ou compostos de coordenação (moléculas constituídas por um ou vários ácidos de Lewis ligados a uma ou várias bases de Lewis.) que reagem tanto com bases quanto com ácidos. Alguns metais, como zinco, cromo, manganês e alumínio, formam óxidos e hidróxidos anfóteros.
No caso dos aminoácidos, eles são denominados assim pois eles podem ter tanto comportamento ácido como comportamento básico, lembrando que, quando falamos em ácido e base, estamos falando a luz da teoria de Brönsted e Lowry.
E uma vez que ácidos e bases se neutralizam formando sal, em um aminoácido ocorrerá uma neutralização intermolecular, formando um sal interno.
Caráter anfótero dos aminoácidos
Os aminoácidos podem ainda sofrer outras divisões, segundo a cadeia lateral ou grupo “R” presente em cada um deles
Aminoácidos apolares
Nesses aminoácidos o grupo “R” ou cadeia lateral é apolar, i.e., são aa hidrofóbicos. Como é o caso da alanina, leucina, valina, cisteína, glicina, prolina, isoleucina, metionanina, triptofano e fenilalanina.
Aminoácidos polares neutros
Nesse grupo de aminoácidos o grupo “R” é uma cadeia lateral polar sem carga elétrica, ou seja, neutra. São hidrofílicos e geralmente contêm hidroxilas, sulfidrilas e aminas. temos como exemplo a glicina, serina, treonina, cisteína, tirosina, asparagina e glutamina.
Aminoácidos polares ácidos
Nesse grupo de aminoácidos encontramos os hidrofílicos e o grupo “R” é uma cadeia lateral com carga negativa, normalmente possuem grupo carboxila, além daquela da estrutura geral. Como exemplo temos o ácido glutâmico e ácido aspártico.
Aminoácidos polares básicos
Nesse grupo estão os aminoácidos hidrofílicos e o grupo “R” é uma cadeia lateral básica, carregada positivamente, possuem grupo amino. Como exemplo temos a histidina, lisina e arginina.
Curiosidades
Como memorizar os aminoácidos
Os alunos de bioquímica costumam se fazer uma das perguntas mais comuns: como memorizar a estrutura de todos os 20 aminoácidos? No entanto, usando algumas dicas, é possível memorizar todos os 20 aminoácidos. E não é tão difícil fazer isso. Você só precisa seguir algumas dicas simples. Primeiro, você precisa generalizar as cadeias laterais de todos esses 20 aminoácidos com base em suas semelhanças.
Olhe para a estrutura da glicina, é o aminoácido mais simples sem carbono quiral. Tenho certeza que você se lembrará da estrutura da glicina para sempre. Portanto, começando com a glicina, você pode desenhar estruturas de todos os aminoácidos, exceto a prolina. Tudo que você precisa para substituir a cadeia lateral por outros grupos.
Substitua um átomo de H no carbono da glicina por um grupo CH3- você obterá alanina. Agora, substitua o átomo H do grupo -CH3 por um grupo fenil, você obterá fenilalanina. Se você adicionar um grupo -OH no anel fenil da fenilalanina na posição para, obterá tirosina. Veja, tão fácil!
Agora vá para a alanina novamente e substitua um átomo de H do grupo -CH3 pelo grupo -OH e você obterá a serina. Tão simples!
Em seguida, substitua um átomo de H do grupo -CH2 da serina pelo grupo -CH3, você obterá treonina. No entanto, se você substituir o átomo O da serina por S, isso lhe dará cisteína. É tão simples. Até agora, você conseguiu se lembrar de estruturas de sete aminoácidos.
Agora, substitua um átomo de H da cadeia lateral da alanina por um grupo –COOH e você obterá ácido aspártico. Isso também é muito simples.
Converter o grupo carboxila do ácido aspártico em um grupo amida lhe dará asparagina. Agora, adicione um grupo -CH2 extra na cadeia lateral do ácido aspártico e obterá o ácido glutâmico. Não é muito simples? Novamente, converta o grupo carboxila da cadeia lateral do ácido glutâmico no grupo amida e você obterá glutamina.
Bem, se você deseja obter metionina, substitua o grupo carboxila da cadeia lateral do ácido glutâmico pelo grupo –S-CH3. Até agora, você memorizou estruturas de 12 aminoácidos e há apenas 8 aminoácidos restantes para memorizar.
Para histidina e triptofano
Agora, você precisa se lembrar das estruturas de um anel imidazol e de um anel indol. No entanto, não existe uma regra de atalho para memorizar esses anéis, mas você sempre pode brincar com esses dois anéis para distingui-los e posicionar o átomo de nitrogênio no anel.
Para histidina, substitua um átomo H da cadeia lateral da alanina por um anel imidazol, você obterá histidina. Da mesma forma, se você remover um átomo de H da cadeia lateral da alanina com um anel indol, obterá triptofano. Isso é tão simples.
Para prolina
No caso da prolina, é preciso entender o que é o grupo imino. Um grupo imino é aquele em que um átomo de nitrogênio está conectado a dois átomos de carbono diferentes, em vez de átomos de hidrogênio. A prolina é o único aminoácido que possui um grupo imino. Isso significa que dois dos átomos de H de seu grupo amino são substituídos por dois átomos de carbono. Outros aminoácidos possuem um grupo amino, mas a prolina possui um grupo imino. Então, para lembrar a estrutura da prolina, conecte o carbono alfa da glicina com o amino N usando um grupo propil. É tão simples. O grupo propil ligado ao amino N e C alfa cria uma estrutura em anel; uma estrutura única da prolina.
Agora, volte para a alanina. Agora, substitua os dois átomos de H da cadeia lateral da alanina por dois grupos CH3, você obterá valina. Se você substituir um átomo de H da cadeia lateral da alanina por um grupo isopropila, obterá leucina. E se você mudar a posição do grupo metil terminal para o carbono beta, obterá isoleucina. Não é tão simples? É muito simples. Agora, faltam apenas dois aminoácidos para os quais você precisa memorizar a estrutura.
Para arginina e lisina
Agora, faltam apenas dois aminoácidos para os quais você precisa memorizar a estrutura e esses aminoácidos são arginina e lisina. Bem, eles também são semelhantes entre si, mas novamente você precisa entender a estrutura de um grupo arginino. Um grupo arginino é semelhante ao da estrutura da ureia. Se você substituir o átomo de oxigênio da ureia por um grupo NH2+, obterá um grupo arginino. É tão simples.
Agora, vá direto ao ponto. Adicione um grupo propil ao carbono alfa da glicina e anexe um grupo arginino no carbono terminal do grupo propil, você obterá arginina. De forma simples, o grupo propil conecta um grupo arginino com o carbono alfa da glicina. Da mesma forma, um grupo butil conecta um grupo amino ao carbono alfa da glicina para formar uma lisina. Você vê, é muito simples construir a estrutura de todos os 20 aminoácidos e memorizar essas estruturas. Boa sorte. (Biochem)
A ligação peptídica é a ligação que une dois ou mais aminoácidos para formar uma molécula. Essa ligação ocorre entre os grupos carboxila, é extremamente forte e libera uma molécula de água.
A molécula aminoácidos formada pode ser chamada de dipeptídeo se tiver apenas dois aminoácidos ou polipeptídeo se for uma macromolécula com mais de três aminoácidos.
As cadeias de polipeptídeos formam as proteínas que podem ter centenas de aminoácidos, a proteína do sangue, a hemoglobina (Hb), por exemplo, possui uma sequência polipeptídica composta por 547 aminoácidos (Flores, 2020).
Estrutura básica dos aminoácidos
Carbono alfa e quando um carbono alfa é quiral
Carbono alfa. Na química orgânica o carbono alfa refere-se ao carbono adjacente ao grupo funcional, o carbono alfa, Cα, é ligado à primeira posição, ou posição alfa, logo após o carbono que possui a função. Por extensão, o segundo carbono é o carbono beta Cβ, e assim por diante.
Carbono quiral é um átomo de carbono que apresenta quatro ligantes diferentes, no caso dos aminoácidos os ligantes são NH3, H, COOH, R; grupo amino, H, grupo carboxila e a cadeia lateral, respectivamente). O carbono quiral é fundamental para identificar a isomeria óptica e o número de isômeros que podem ser formados. Denomina-se de carbono quiral, ou carbono assimétrico, o carbono de uma cadeia que apresenta quatro ligantes diferentes.
Dos vinte (20) aminoácidos mais comuns existentes o único aminoácido que não apresenta carbono quiral é a glicina (Gli), pois apresenta três radicais diferentes (N2H, COOH, H, H), havendo dois radicais hidrogênio.
Estrutura básica dos aminoácidos.
Todo aminoácido apresenta um carbono central (carbono alfa) onde estão ligados: um grupo amina (NH2), uma cadeia lateral representado pelo R (radical, que diferencia cada um dos 20 aa), também chamado de grupo variável, e o grupo carboxila (COOH). A cadeia lateral ou radical determina as propriedades do aa, pois o R pode ser polar (hidrofílico), negativo (carregado negativamente), não polar (apolar, hidrofóbico) ou carregado positivamente; (hidrofóbico, ácido, básico, positivo, negativo, hidrofílico).
Os aminoácidos se ligam entre si através da reação
chamada de reação de condensação, resultando numa molécula de água.
A reação acontece com o OH do grupo carboxílico e o H do grupo amina, formando uma ligação peptídica, que nada mais é do que uma ligação covalente.
Lista dos vinte aminoácidos e sua estrutura
Observe que o R (cadeia lateral ou radical)
varia de aminoácido para aminoácido
As proteínas são sintetizadas utilizando as informações contidas no material genético, armazenadas no DNA, transcritas e transportadas no mRNA para o citoplasma onde o mRNA será traduzido pelos ribossomos.
O processo de síntese protéica, também chamado de tradução, ocorre nos ribossomos, sendo necessário além do mRNA, o tRNA, aminoácidos e ATP.
Assim como os carboidratos polissacarídeos, que são formados por unidades menores de monossacarídeos, as proteínas são polímeros formados por unidades chamadas de aminoácidos.
Os aminoácidos são compostos orgânicos constituídos de um carbono central (em que estão ligados todos os demais grupos) chamado de carbono alfa. Em uma extremidade do aminoácido há um grupamento amina (NH2) chamado de porção N-terminal. Em outra extremidade há um grupamento carboxila (COOH) chamado de porção C-terminal e uma extremidade chamada de radical R, que varia de um aminoácido para outro e é utilizado como forma de identificação de cada um dos resíduos de aminoácidos.
Para completar as quatro ligações que o carbono precisa ter para ficar “estável” a maioria dos aminoácidos possui na quarta extremidade um hidrogênio (H).
A estrutura tridimensional de cada proteína é determinada pela sequência de aminoácidos que formam cada polipeptídeo. Assim, podemos distinguir quatro níveis estruturais em proteínas.
Estrutura primária
Não é propriamente uma estrutura, uma vez que a ordem dos animoácidos na cadeia polipeptídica. Forma-se quando ocorre a ligação entre os monnômeros (resíduos de aa) da cadeia principal polipeptídica. As junções desses elementos são por meio de ligações peptídicas (ligações covalentes).
A estrutura primária é dada pela sequência linear dos aminoácidos, é esta sequência de aminoácidos, é determinada pelo gene no DNA. Dependendo do tipo de aminoácido termos diferentes estruturas secundária e terciária.
Estrutura secundária
A estrutura secundária é resultante da interação entre os aminoácidos dentro da cadeia ou na cadeia polipeptídica, e vai depender dos radicais que cada um dos aminoácido possuir. Pode-se observar que as cadeias são torcidas, dobradas ou enroladas sobre elas mesmas. As mais comuns são a alpha-hélice e a beta-folha (ou beta-hélice, ou beta-lâminas). Essas interações entre os aminoácidos são mantidas por ligações pontes de hidrogênio. A alfa-hélice é pelo estabelecimento de interações do tipo pontes de hidrogênio entre o H do grupo amino (-NH) e o O da carbonila (C=O). Além dessas interações que formam a alpha-hélice e as beta-folha, ainda ocorrem as "beta-turn" (voltas-beta) e ômega-loop.
Alpha-hélice
Beta-plated sheet ou Beta-folha ou folha pregueada
Estrutura terciária
Corresponde à forma adquirida pelo polipeptídeo depois da interação entre as cadeias laterais dos aminoácidos que constituem essa proteína.
Estrutura quaternária
Podemos notar uma estrutura quaternária quando houver associação de duas ou mais cadeias polipeptídicas. A estrutura quaternária é mantida principalmente por ligações iônicas, pontes de hidrogênio e por ligações do tipo hidrofóbico.
Estruturas encontradas nas proteínas e o que as mantém
As proteínas também podem ser classificadas em simples, conjugadas e derivadas.
Proteínas simples: formadas apenas por aminoácidos.
Proteínas conjugadas: quando sofrem hidrólise, liberam aminoácidos e um radical não peptídico. Esse radical é denominado de grupo prostético.
Proteínas conjugadas e seu grupo prostético
Grupo prostético é a designação de uma substância não protéica quando está combinada com uma proteína. não deve ser confundido com cofator que atuam no sitio alostérico das enzimas. Muitos pigmentos respiratórios são proteínas conjugadas. Os carboidratos e lipídios nas glicoproteínas e lipoproteínas são grupos prostéticos de suas proteínas, da mesma forma, o DNA (ácido desoxirribonucleico) é o grupo prostético das histonas na cromatina.
Proteínas derivadas: não são encontradas na natureza e são obtidas pela degradação, por meio da ação de ácidos, bases ou enzimas, de proteínas simples ou conjugadas.
DE ACORDO COM A FUNÇÃO PODEMOS DIVIDIR AS PROTEINAS EM ALGUMAS CLASSES COMO ESTRUTURAL, TRANSPORTADORA, NUTRITIVA, DEFESA E CATALIZADORA.
TODAS SÃO IGUALMENTE IMPORTANTES MAS DUAS DELAS DESEMPENHAM FUNÇÕES IMPRESCINDÍVEIS NOS ORGANISMOS: PROTEÍNAS ESTRUTURAIS E PROTEÍNAS ENZIMÁTICAS OU CATALÍTICAS.
Proteínas estruturais
São possuem com função constitutiva e construtiva em diversas células e tecidos. Ex.: Queratina, Colágeno.
O colágeno é a proteína mais comum nos metazoários, constituindo o arcabouço extracelular para todos os organismos multicelulares.
Sem o colágeno, o ser humano seria reduzido a um aglomerado de células, semelhante ao “Blob” (o “horror gelatinoso do espaço” famoso, em filme da década de 1950, que teve um remake em 1988 como "A bolha assassina"), interconectado por alguns neurônios. (1)
The blob. A bolha assassina.
The Blob é um filme norte-americano de terror e ficção científica de 1958, dirigido por Irvin Yeaworth, e escrito por Kay Linaker e Theodore Simonson.
Um remake foi produzido 1988 com o mesmo nome, que foi co-escrito e dirigido por Chuck Russell.
Atualmente, são conhecidos 27 tipos diferentes de colágenos, codificados por 41 genes dispersos em pelo menos 14 cromossomos.
Cada colágeno é composto de três cadeias que formam um trímero na forma de hélice tripla. O polipeptídeo é caracterizado por uma sequência que se repete, na qual a glicina está na terceira posição (Gly-X-Y, em que X e Y podem ser qualquer outro aminoácido que não cisteína ou triptofano) e contém os aminoácidos especializados 4-hidroxiprolina e hidroxilisina.
Os resíduos de prolina na posição Y são hidroxilados para produzir hidroxiprolina, que estabiliza a hélice tripla.
Os tipos I, II, III, V e XI são os colágenos fibrilares, nos quais o domínio da tripla hélice é ininterrupto por mais de 1.000 resíduos; essas proteínas são encontradas nas estruturas fibrilares extracelulares.
Os colágenos tipo IV possuem domínios longos da tripla hélice, porém interrompidos, e formam bainhas em vez de fibrilas, constituindo os principais componentes da membrana basal, em associação com a laminina.
As lamininas são uma família de (15 tipos de lamininas) glicoproteinas de fundamental importância no desenvolvimento embrionário, além de terem um papel importante em processos de diferenciação, migração e adesão celular.Além disso, dirigem o crescimento celular e são um fator de sobrevivência para diferentes tipos celulares. A laminina é encontrada, em grande parte, nas membranas basais, formando uma rede proteica que organiza a própria formação dessa matriz extracelular especializada. (A, B)
Ilustração de uma proteína estrutural chamada Laminina.
Laminina
Essa molécula é constituída por três polipeptídeos em forma de cruz e também apresenta porções que se ligam ao colágeno tipo IV, ao heparan sulfatado e a receptores celulares de laminina, formando assim pontes que ligam as células à matriz. Como o colágeno IV e o heparan sulfatado são os principais componentes das lâminas basais, a laminina serve de ponte entre as células e essas lâminas.
A ligação do epitélio à lâmina basal é coordenada pela ação dessas proteínas reguladoras. Há momentos em que esta ligação pode ser desfeita, como quando uma célula do estrato germinativo está em divisão, seu citoesqueleto é redirecionado aos eventos cromossômicos, quando a célula precisa realizar movimentos livres para a citocinese, ou quando há condições anormais, como a tração do epitélio causando ruptura desta ancoragem. O espaço gerado por seu desprendimento é rapidamente preenchido por exsudato plasmático do conjuntivo, formando uma bolha no epitélio.
Um outro colágeno com domínio de tripla hélice longo e interrompido (tipo VII) forma as fibrilas de ancoragem entre algumas estruturas epiteliais e mesenquimais, como na epiderme e derme. Outros colágenos, ainda, são transmembrana e podem auxiliar também a ancoragem de estruturas epidérmicas e dérmicas.
Proteínas enzimáticas ou catalíticas
São uma classes protéica que possui função catalítica, sendo responsáveis por acelerar reações bioquímicas. Ex.: alfa-amilase, esterase, peroxidase.
Proteínas enzimáticas e suas características
As proteínas enzimáticas favorecem as reações biológicas
e isso é observado pela diminuição da energia de ativação.
Proteínas transportadoras
Carregam substâncias específicas de um ponto a outro no interior celular.
Ex.: Hemoglobina (anelídeos e vertebrados), Hemocianina (invertebrados)
Proteínas mecânicas
Estão relacionadas aos processos de mobilidade, contração e relaxamento celular e muscular. Ex.: Actina, Miosina.
Proteínas nutritivas
Nutrem ou armazenam conteúdo nutritivo em células específicas. Ex.: Albumina (ovos), Caseína (leite), ovoalbuminas e ovovitelinas.
Proteínas de defesa
São os anticorpos que protegem o organismo contra agentes estranhos. Ex.: Imunoglobulinas, Lisinas, Aglutininas
As proteínas formam uma estrutura tridimensional, a qual pode ser desfeita caso alterações no ambiente ocorram. Dizemos que ocorreu a desnaturação de uma proteína quando ela se desfaz de sua estrutura secundária, terciária e quaternária e perde a sua forma original, tornando-se inativa.
Desta forma ao perder sua estrutura a proteína perde sua conformação específica, perdendo também sua capacidade de exercer as funções catalíticas no organismo.
Um caso emblemático em nossa espécie ocorre com a febre (a elevação rápida da temperatura corporal devido a infecção; quando a temperatura sobe, afastando-se de seus níveis seguros, as moléculas protéicas podem desnaturar. A febre tem sido considerada uma tentativa do organismo de desnaturar as proteínas virais ou bacterianas, todavia, esse mecanismo funciona como uma faca de dois gumes. Desnatura as proteínas invasoras mas também as do nosso corpo. Por isso, devemos combater a causa da febre, para evitar que nosso corpo eleve a temperatura para tentar livrar-se do invasor.
A febre é um aviso do corpo de que alguma coisa errada; isso acontece no caso de invasão por vírus, bactérias e outras entidades biológicas.
O maior perigo da febre é a desnaturação de proteínas essenciais ao nosso corpo.
A febre funciona como uma tentativa de desnaturar proteínas e outros elementos de invasores virais, bacterianos, entre outros. Com o aumento da temperatura corporal, fica mais difícil para que os parasitas consigam funcionar com total liberdade e de maneira eficaz.
Entretanto uma febre descontrolada pode acabar desnaturando macromoléculas biológicas fundamentais para o funcionamento do organismo, além dos vírus e bactérias. Por isso, é importante sempre tentar diminuir ou baixar a temperatura corporal por meio de banho e medicamentos específicos, os antipiréticos, que trazem a temperatura para seu nível normal.
Nosso corpo possui uma temperatura média que varia entre 35.5 até 36.5 graus célsius. Uma temperatura maior que essa, já pode ser considerado em estado febril.
Pode acontecer que a temperatura alcance 40 graus célsius, o que é muito perigoso, já que essa temperatura se mantida por um longo período certamente trará prejuízos para nosso organismo.
Proteínas globulares e fibrosas
As proteínas podem ser classificadas em globulares e fibrosas. As proteínas globulares são aquelas que possuem formas esféricas e são dobradas várias vezes. As proteínas fibrosas apresentam formato de fibra alongada. Quando comparamos as proteínas globulares com as fibrosas, percebemos que essas últimas são menos compactas e mais longas.
Entendemos por desnaturação protéica o processo de alteração da forma de uma proteína sem que a alteração na sua sequência básica de aminoácidos.
Essa desnaturação pode ocorrer com a exposição da proteína a “agentes desnaturantes” como temperatura elevada, alterações do pH, radiação ultravioleta, entre outros.
As proteínas, para poderem atuar adequadamente, dependem de condições físico-químicas bem específicas. Uma proteína que atua em pH ácido não consegue desempenhar sua função em pH neutro ou básico, pois a presença de íons H+ no meio altera sua estrutura secundária, terciária e quaternária, impedindo que os sitio ativo e alostérico se formem e assim a proteína não consegue executar sua função.
O que é ocorre é que as mudanças nas características físico-químicas do meio em que a proteína está, desestabiliza suas ligações mais “fracas”, como as ligações de hidrogênio e as ligações dissulfeto presentes nas estruturas secundárias e terciárias, respectivamente, inativando a proteína.
Fatores que influenciam o funcionamento das enzimas
A desnaturação protéica é a perda da estrutura quaternária, terciárias e secundárias de uma cadeia polipeptídica, fazendo com que a proteína permaneça apenas em sua estrutura primária, ou seja, mantendo apenas as ligações peptídicas que ordenam seus aminoácidos na cadeia. Na desnaturação a sequência de aminoácidos não se altera e nenhuma ligação peptídica é rompida.
Temperatura, pH e desnaturação
Uma vez retirada a proteína dessas condições desnaturantes, elas podem reassumir sua conformação nativa pelo processo de renaturação. (Manual do Enem)
01) Leia o texto a seguir, escrito por Jacob Berzelius, em 1828:
“Existem razões para supor que, nos animais e nas plantas, ocorrem milhares de processos catalíticos nos líquidos do corpo e nos tecidos. Tudo indica que, no futuro, descobriremos que a capacidade de os organismos vivos produzirem os mais variados tipos de compostos químicos reside no poder catalítico de seus tecidos.”
A previsão de Berzelius estava correta, e hoje sabemos que o “poder catalítico” mencionado no texto deve-se a uma espécie de composto muito importante.
a) que composto é esse? E como esse "composto" esta estruturado? Explique.
02) (UFSCar) Durante um estudo sobre a digestão humana, foi realizado o seguinte experimento:
É correto afirmar que a digestão da carne ocorreu apenas:
a) No tubo I.
b) No tubo II.
c) No tubo III.
d) Nos tubos I e II.
e) Nos tubos II e III.
03) (UFSCAR) Ao preparar um lanche foram usados pão francês, maionese, filé mignon, queijo e alface. O primeiro desses ingredientes a sofrer ação de enzimas digestivas é:
a) A alface.
b) O queijo.
c) A maionese.
d) O filé mignon.
e) O pão francês.
justifique sua resposta.
04) (UNIVASF) No quadro abaixo, são mostradas algumas reações que ocorrem no trato digestivo humano. Assinale a alternativa que indica, respectivamente, enzimas que atuam em 1, 2 e 3.
Quando foi encontrada por Layard, essa peça oval de quartzo polido ou cristal de rocha foi imediatamente identificada como uma lente e passou a ser conhecida como 'lente Nimrud'. Certamente poderia ter sido usado como uma lupa tosca, com uma distância focal de 12 centímetros da superfície plana. Ao longo dos anos, ela foi examinada por vários oftalmologistas (por exemplo, Gasson 1972), muitos dos quais acreditam que foi deliberadamente fabricada como uma lente. No entanto, embora esta peça de cristal de rocha tenha sido cuidadosamente lixada e polida e, sem dúvida, tenha propriedades ópticas, elas provavelmente são acidentais. Não há evidência de que os assírios usavam lentes, seja para aumentar ou para fazer fogo, e é muito mais provável que se trate de um pedaço de quartzo para incrustação, talvez para móveis. Isso é corroborado pela afirmação de Layard de que esse objeto "foi enterrado sob uma pilha de fragmentos de um belo vidro opaco azul, aparentemente o esmalte de algum objeto de marfim ou madeira, que pereceu" (Layard 1853: 198).
Os romanos já usavam lentes biconvexas. Existem relatos de que Nero, imperador romano (37 - 68 d.C.), assistia combates de gladiadores com o auxílio de uma esmeralda talhada. O que nos leva a supor que era conhecida, em alguma medida, a propriedade das lentes para correção da miopia.
Porém as lentes passaram a ser realmente conhecidas e utilizadas por volta do ano 1280 na Itália, com a invenção dos óculos.
Eventos importantes na historia da
citologia ou biologia celular
Hans Lipperhey e Zacharias Jansen
Hans Lipperhey (1570 - 1619) germano-holandês , também conhecido como Johann Lippershey, nasceu em Wesel, agora no oeste da Alemanha era um fabricante e polidor de lentes para óculos . É comumente associado à invenção do telescópio, pois foi o primeiro a tentar obter a patente do mesmo.
Hans Lippershey é conhecido pelo mais antigo registro escrito de um telescópio refrator, por uma patente que ele tentou registrar em 1608.
Seu trabalho com dispositivos ópticos cresceu fora de seu trabalho como fabricante de óculos, uma indústria que começou em Veneza e Florença no século XIII, (1200) e mais tarde se expandiu para a Holanda e Alemanha.
Lipperhey solicitou aos Estados Gerais da Holanda em 2 de outubro de 1608 a patente de seu instrumento "For seeing things far away as if they were nearby" (para ver as coisas distantes como se estivessem perto), algumas semanas antes da patente de outro fabricante de instrumentos holandês, a de Jacob Metius. Lipperhey não conseguiu receber uma patente, pois a mesma reivindicação de invenção também havia sido feita por outros fabricantes de óculos, mas ele foi generosamente recompensado pelo governo holandês por cópias de seu projeto.
O pedido de patente de Lipperhey foi mencionado no final de um relatório diplomático sobre uma embaixada do Reino de Sião na Holanda, enviado pelo rei siamês Ekathotsarot: Ambassades du Roy de Siam enviado à l'Excelency du Prince Maurice, arrivé à La Haye le 10 de setembro 1608 (Embaixada do Rei do Sião enviada a Sua Excelência o Príncipe Maurício, chegou a Haia em 10 de setembro de 1608). Este relatório foi publicado em outubro de 1608 e distribuído por toda a Europa, levando a experimentos de outros cientistas, como o italiano Paolo Sarpi, que recebeu o relatório em novembro, o inglês Thomas Harriot, que usava um telescópio de seis potências no verão de 1609, e Galileo Galilei, que melhorou o dispositivo.
Existem muitas histórias sobre como Lipperhey chegou a sua invenção. Uma das versões mostra Lipperhey observando duas crianças brincando com lentes em sua loja e comentando como elas poderiam fazer um cata-vento (weather-vane) distante parecer mais próximo ao olhá-lo através de duas lentes. Outras histórias têm o aprendiz de Lipperhey tendo a ideia ou Lipperhey copiando a descoberta de outra pessoa.
O instrumento original de Lipperhey consistia em duas lentes convexas com uma imagem invertida ou uma objetiva convexa e uma lente ocular côncava para que tivesse uma imagem direita vertical (upright image). Este "vidro de perspectiva holandesa" (o nome "telescópio" só seria cunhado três anos depois por Giovanni Demisiani) tinha uma ampliação de três vezes (ou 3X).
A cratera lunar Lipperhey, o planetoide 31338 Lipperhey e o exoplaneta Lipperhey (55 Cancri d) receberam seu nome.
Zacharias Janssen (1585 - 1632) nasceu em na cidade de Haia (The Hague), Holanda (Países Baixos), filho de Hans Martens Janssen (muito provavelmente vendedor ambulante)(1) e sua esposa Maeyken Meertens (ambos da Bélgica), e cresceu na cidade Middelburg. Zacharias Janssen tornou-se fabricante de lentes de óculos, e Junto ao também inventor germano-holandês Hans Lippershey, Zacharias Janssen é creditado como inventor do microscópio composto com duas lentes no início de 1590 séc. XVI e do telescópio.
Hans Lippershey (1570 -1619
(Fonte: Pierre Borel - De vero telescopii inventore 2, 3 )
Zacharias Jansen (1585 - 1632)
(Fonte: Pierre Borel, De vero telescopii inventore, Public Domain).
Reproduction of an optical device that Zacharias Snijder in 1841 claimed was an early telescope built by Jansen. Dutch biologist and naturalist Pieter Harting claimed in 1858 that this was an early microscope which he also attributed to Janssen, perpetuating the Janssen claim to both devices. Its actual function and creator has been disputed. (4)
O microscópio de Zacharias e Lippershey era construído usando duas lentes convexas com uma imagem invertida ou uma objetiva convexa e uma lente ocular côncava para que tivesse uma imagem direita vertical (upright image). Um o objeto colocado perto do final do tubo parece então ser muito ampliado, maior do que qualquer lupa simples poderia alcançar. A princípio era tratado como um brinquedo pelos nobre da realeza europeia.
Robert Hooke indica com ou termo célula, as secções discretas que identifica numa amostra de cortiça observada num dos primeiros microscópios. Dez anos depois, publica Micrographia, reconhecendo que não viu as células, agora mortas, mas as suas paredes celulares numa lâmina cortiça.
1674
Antoni van Leeuwenhoek identifica o protozoário, que ele chama de animálculos.
Ele também identificou bactérias. Suas descobertas, fragmentárias e pouco conhecidas devido serem escritas e publicadas em holandês e não inglês, foram possíveis graças à sua experiência em construir um sistema de ampliação com lentes polidas.
1833
Robert Brown descreve o núcleo da célula de uma planta de orquídea.
1838
Matthias Schleiden e Theodor Schwann propõem a teoria celular, argumentando que as unidades básicas dos seres vivos são células, contendo um núcleo. É o nascimento formal da citologia ou Biologia celular.
1852
Albert von Kölliker, biólogo, anatomista e fisiologista suíço, descreve em 1857 as mitocôndrias como grânulos em em células musculares.
Albert von Kölliker (1817 - 1905)
1858
Rudolf Virchow, medico, antropólogo e político liberal é considerado o pai da patologia moderna e da medicina social. Virchow argumenta que todo ser vivo multicelular é a soma de um grande número de unidades únicas totalmente viáveis, células, provenientes de outras células pré-existentes, ele cunha a famosa frase "Omnis cellula ex cellula", do latim: "Toda célula de (provém) uma célula". Partindo da mesma suposição, em Zellpatologie argumenta que as patologias do organismo se originam de patologias celulares.
1879
Walther Flemming, biólogo alemão, considerado o fundador da citogenética. Usando corandes (anilina) descobriu e nomeou a cromatina (liga-se a corantes bsófílicos) e que esta estava correlacionada com os cromossomos (corpos coloridos) descreve detalhadamente o comportamento dos cromossomos durante a mitose das células animais.
Wather Flemming (1843 - 1905)
Cromossomos politênicos em uma célula da glândula salivar de Chironimus, um dos mais de 100 desenhos do livro de Flemming Zellsubstanz, Kern und Zelltheilung, 1885.
(Fonte: Walther Flemming. Professor der Anatomie in Kiel. Zellsubstanz, Kern und Zelltheilung. Leipzig, Verlag von F.C.W. Vogel. Public Domain).
1881
Anders Retzius descreve em detalhes muitos tecidos de animais. Nas duas décadas seguintes, junto com Santiago Ramón y Cajal e outros histologistas, ele desenvolveu várias técnicas de coloração, estabelecendo as bases da anatomia microscópica moderna.
1882
Robert Koch usa corantes contendo anilina para realçar as células dos microrganismos, identificando os responsáveis pelo cólera e pela tuberculose. Outros bacteriologistas, como Edwin Klebs e Louis Pasteur, posteriormente identificam os organismos responsáveis por muitas outras patologias.
Robert Koch (1843 - 1910)
1898
Camillo Golgi, medico e histologista italiano, identifica e descreve o sistema intercelular conhecido hoje como aparelho de Golgi, colorindo as células com nitrato de prata.
Camillo Golgi (1843 - 1926)
1912
Jacques Loeb retira óvulos de ouriços-do-mar, e induz quimicamente sua embriogênese, para observar os estágios da embriogênese.
1931
Ernst Augost Friedrich Ruska, o engenheiro e físico alemão inventa o primeiro microscópio eletrônico de transmissão. Esse equipamento possui um poder de resolução astronômico quando comparado com o microscópio ótico. (ver tb: Sites)
Em 9 de março de 1931, o engenheiro eletrico e físico alemão Ernst Ruska e seus colaboradores apresentam o primeiro microscópio eletrônico de transmissão, a projeção deste aparelho e a sua importância foi tão grande que 55 anos após (1986) Ruska recebeu o Prêmio Nobel de física. A microscopia eletrônica de varredura teve seu início com o trabalho de M. Knoll (1935), descrevendo a concepção do MEV. Em 1938 von Ardenne construiu o primeiro microscópio eletrônico de varredura adaptando bobinas de varredura ao microscópio eletrônico de transmissão. O primeiro microscópio eletrônico de varredura para observação de amostras espessas foi construído em 1942 nos laboratórios da RCA usando o detector de elétrons secundários para obter a imagem. Em 1939 a empresa Siemens produz o primeiro microscópio de transmissão comercial. Em 1957, pela primeira vez Robertson observa e descreve a estrutura trilaminar de membrana. Em 1965 foi produzido comercialmente pela empresa Cambridge Scientific Instrument o primeiro MEV. Durante este período várias técnicas, reagentes (criofratura, sombreamento de metal, fixadores, resinas de epóxi araldite etc.) e aparelhos como ultramicrótomo foram desenvolvidos para a preparação de amostras para visualização ao ME.(6, 7, 8, 9)
A) Esquema mostrando como a luz é ampliada ao passar por um conjunto de lentes no microscópio. B) Esquema de um microscópio óptico; C) Primeiro microscópio eletrônico de transmissão construido por Ernst Ruska em 1931;
A maioria das células tem entre 1μm e 100μm de diâmetro por isso são visíveis apenas no microscópio.
A razão da área da superfície pelo volume é crítico para a célula. Quando aumenta a área da superfície da célula, esta (a área) aumenta por um fator igual a n2 enquanto que o volume aumenta por um fator igual a n3. A área aumenta ao quadrado e o volume aumenta ao cubo. Assim, as células pequenas tem maior área superficial do que volume.
CÉLULA PROCARIOTA (em construção)
Os primeiros estudos morfológicos usando microscopia ótica e até mesmo as observações iniciais feitas com microscópios eletrônicos de transmissão indicaram grandes diferenças na organização das células procarióticas e eucarióticas.
Enquanto as células eucarióticas foram caracterizadas como exibindo organização interna associada a várias organelas com com estrutura membranosa, as bactérias exibiram uma organização interna homogênea.
Embora essa ideia tenha predominado por muitos anos, a análise detalhada da organização estrutural das células procarióticas, especialmente usando abordagens estruturais modernas em associação com análises genômicas e proteômicas (estudo das proteínas), indicou que essa visão está incorreta.
De fato, as informações disponíveis agora apontam para a existência de um conjunto de estruturas proteicas complexas localizadas em regiões bem definidas da célula procariótica, bem como a presença de várias organelas delimitadas por membrana, indicando a existência de subcompartimentos nessas células. A existência de um citoesqueleto complexo formado por proteínas semelhantes às envolvidas na montagem de microtúbulos, microfilamentos e filamentos intermediários indica ainda que algumas características estruturais que geralmente estão associadas a processos fisiológicos fundamentais, como manutenção da forma celular, divisão celular e locomoção celular, têm mantido ao longo do processo evolutivo.
Além disso, foi demonstrado que processos complexos, como glicosilação de proteínas (Benz & Schmidt 2002) e adesão célula-célula, que antes eram reconhecidos apenas em células eucarióticas, também são comuns em bactérias que estão comprometidas umas com as outras e se desenvolvem em multicelulares estruturas (Keim et al. 2004).
Cápsula: reveste a célula externamente;
Citoplasma: substância gelatinosa responsável por manter o formato da célula;
DNA: carrega as informações genéticas da célula;
Flagelo: possibilita a locomoção da célula;
Membrana plasmática: controla a troca de substâncias da célula com o meio externo;
Parede celular: confere forma à célula;
Pilus: possibilita a fixação da bactéria ao meio;
Ribossomo: estrutura responsável pela produção de proteínas.
PROCARIOTA
EUCARIOTA
NUCLEIÓIDE mesossoma
ancora o DNA
NÚCLEO Carioteca reveste
o genoma (todos os cromossomos)
DNA circular no citosol
DNA linear dentro do
núcleo
Apenas UM cromossoma
(algumas bactérias tem dois: Vibrio
cholerae
Mais de um crossomo
Moléculas lineares de
DNA individualizadas
Sem Histonas
Molécula de DNA se
enovela com as histonas
A transcrição do DNA e
a tradução do RNA ocorrem simultaneamente no citosol
Transcrição do DNA no
núcleo
Tradução do RNA no
citosol
Genoma haplóide, i.e.,
somente uma cópia de cada gene
Genoma diploide, i.e.,
cada gene tem duas cópias; alguns podem ser haplóides parte do tempo, e
muitas espécies podem ser haplóide
Presença de um ou mais
plasmídeos (moléculas de DNA fora do cromossomo)
Sem plasmídeos
Uma única origem para
a replicação do genoma
Múltiplas origens para replicação
Genoma compacto, pouco
repetitivo
Geralmente grandes
quantidades de DNA não codificante e repetitivo
Genes sem introns
Genes com introns
Sedimentação do
ribossomo mais lenta (70S)
Sedimentação dos
ribossomos mais lenta (80S)
Ribossomos pequenos
Grandes ribossomos no
citosol, e carioteca, organelas
pequenas em organelas.
Sem organelas membranosas
Muitas organelas
delimitadas por membrana.
Membrana plasmática sem
receptores
Membrana plasmática
com receptores, esteróis e carboidratos)
Parede celular
quimicamente complexa, com peptídeo glicano (Mycoplasma sp sem parede celular)
Parede celular quimicamente
simples, Celulose em plantas e quitina em fungi
Flagelo se presente
simples construído de duas proteínas
Apresenta ribossomos os quais secretam glicoproteínas (proteínas ligadas por ligações covalentes a carboidratos).
Distribui vesículas de transporte, proteínas rodeadas de membrana.
É a fábrica de membranas da célula.
Núcleo celular
Ribossomos
Complexo de Golgi ou Aparato de Golgi ou Golgiense
Consiste de sacos de membranas achatadas e empilhados chamados de cisternas cujas regiões laterais são dilatadas, e tem como funções:
Modificar os produtos do Reticulo endoplasmático
Fabricar certas macromoléculas
Classifica e empacota materiais em vesículas de transporte.
O complexo golgiense é uma estrutura polarizada, apresentando duas faces: a face cis e a face trans:
A face cis, ou face de formação, é uma superfície convexa e o local responsável por receber as vesículas provenientes do retículo endoplasmático, sendo o local mais próximo do retículo. Nela, as vesículas provenientes do retículo endoplasmático fundem sua membrana e liberam seu conteúdo.
A face trans, ou face de maturação, por sua vez, é a face côncava e é a responsável por gerar vesículas que partem do complexo, indo para outras partes da célula. Vale destacar ainda que, entre as duas faces, há as chamadas cisternas medianas ou mediais.
O número de cisternas presentes em uma pilha e o número de pilhas encontrados em uma célula variam muito de uma célula para outra. Essa organela é bastante desenvolvida em células especializadas na secreção de substâncias, como é o caso das células caliciformes localizadas no intestino.
Lisossomos
O lisossomo é uma vesícula ou saco membranoso com enzimas hidrolíticas que podem digerir macromoléculas.
As enzimas dos lisossomos podem hidrolisar proteínas, gorduras, polissacarídeos e ácidos nucléicos. Enzimas presentes nos lisossomos: lipases, proteases, glicosidases, fostatases, nucleases, sulfatases.
Sua função é a digestão intracelular.
Os lisossomos são produzidos no Complexo de Golgi onde são formadas vesículas que se soltam originando os lisossomos primários (vesículas cheias de enzimas). Esses lisossomos ficam no citoplasma onde atuam, até que a célula realize endocitose (fagocitose ou pinocitose) e englobe alguma partícula externa (sólida ou líquida). Nesse processo, a partícula é interiorizada dentro de uma vesícula, chamada endossomo, que se funde com o lisossomo primário formando o lisossomo secundário, que é uma espécie de vacúolo digestivo.
Quando a célula necessita digerir substâncias vindas do meio externo, ela realiza fagocitose. Por exemplo, no caso das células do sistema imunitário humano que atacam células inimigas chamadas antígenos.
Uma célula inimiga que pode ser uma bactéria, por exemplo é capturada por uma célula APC (Antigen Presenting Cell): célula apresentadora de antígeno ou célula acessória), que pode ser um macrófago ou um linfócito através da fagocitose. Para que ocorra fagocitose é necessário reconhecimento. A bactéria é então reconhecida, ligada à e envolvida pela membrana plasmática do macrófago e forma uma vesícula chamada fagossomo. No citoplasma da célula, o fagossomo se funde ao lisossomo, e em seguida, as enzimas digestivas contidas no lisossomo começam a agir. O microrganismo ou o corpo invasor é digerido ou lisado ou quebrado em partes menores e eliminado para fora da célula como moléculas residuais. Os lisossomos também são responsáveis pela destruição de organelas envelhecidas dentro da célula, processo que chamamos de autofagia. Através desse processo digere algumas das suas organelas que já não funcionam mais. Isso também pode acontecer em situações com poucos nutrientes, em que a célula realiza a autofagia para manter a homeostase (equilíbrio interno).
Vacúolos
Existem três tipos de vacúolos: os vacúolos digestivos, os vacúolos contráteis e os vacúolos de armazenamento.
A função de cada tipo de vacúolo vai depender do organismo em que se encontram.
Os vacúolos são estruturas citoplasmáticas com tamanhos variados e revestidos por membrana, formados a partir do retículo endoplasmático ou do complexo de Golgi.
São encontrados em protoctistas nos protozoa (vacúolos digestivos, contráteis e armazenamento) e algas (unicelulares e pluricelulares), nas células vegetais, fungos e animais.
Nas células animais os vacúolos são pequenos e em maior número do que nas células vegetais, que possuem um único que às vezes ocupa metade do volume celular.
Vacúolos digestivos
Os digestivos, típicos de células fagocitárias, estão associados à atuação de enzimas lisossômicas, formando outros vacúolos derivados a esse processo: vacúolos primários e secundários, ou também chamados de vacúolos digestivos e residuais.
Vacúolos contráteis ou pulsáteis
Os contráteis, ou também com função pulsátil, são característicos de alguns protozoários como Paramecium por ex. que são responsáveis pelo equilíbrio osmótico, eliminando o excesso de água.
Vacúolo de armazenamento
Os vacúolos de armazenamento, são encontrados sobretudo nos vegetais, ocupam um volume celular significativo. Dependendo da espécie considerada, armazenam variadas substâncias: carboidratos (amiloplastos, plastos de reserva nutritiva nas raízes), proteínas (proteoplastos, plastos de reserva protéica em sementes), alguns pigmentos de pétalas, semente e folhas (antocianinas) e até toxinas (nicotina e tanino), substâncias secundárias de defesa contra predadores herbívoros (deterrentes).
Os vacúolos são organelas citoplasmáticas com especialização evolutiva bastante diversificada.
Juntamente com os plastídios ou plastos e a parede celular, o vacúolo é uma das três estruturas que distinguem as células vegetais das células animais.
Nos vegetais os vacúolos são regiões dentro da célula delimitadas por uma única membrana e repletas com um líquido denominado suco celular.
A membrana que circunda o vacúolo é denominada tonoplasto.
O principal conteúdo do suco vacuolar é a água, os outros componentes variam de acordo com o tipo de célula, órgão ou especie de planta e de seu estágio de desenvolvimento ontogenético e fisiológico.
Além dos íons inorgânicos como cálcio, potássio, cloro, sódio e fósforo, o vacúolo comumente contém pigmentos, proteínas, açúcares, ácidos orgânicos e aminoácidos.
Algumas substâncias no vacúolo podem se solidificar por ex.: taninos e proteínas ou até cristalizar-se como ocorre com os cristais de oxalato de cálcio.
Cristais de oxalato de cálcio, que podem assumir várias formas diferentes, são especialmente comuns. O suco vacuolar usualmente é levemente ácido, mas em alguns casos, como aqueles dos vacúolos das células dos frutos de Citrus sp., é muito ácido, conferindo o gosto azedo e ácido ao fruto.
A. Células com vacúolos contendo substâncias fenólicas em
Aleuroma em Ricinus communis L. (1753) (?). Aleurona (do grego aleuron, farinha) é uma proteína encontrada em grânulos principalmente em endosperma de sementes e em tubérculos em maturação. As proteínas de Aleurona podem aparecer em duas características morfológicas diferentes, homogêneas e heterogêneas. A aleurona homogênea consiste em corpos proteicos semelhantes (por exemplo, em Phaseolus vulgaris), enquanto a aleurona heterogênea consiste em grânulos de diferentes formas e tipos de proteínas cobertas por uma membrana (por exemplo, em Ricinus communis).. (Apezzato-Guerreiro, 2003)
A mitocôndria do grego: μίτος = mitos: fio/linha e χονδρίοv = chondrion: grânulo; é a organela mais importante pois é responsável pela respiração celular, um processo metabólico que produz energia na forma de ATP.
É abastecida pela célula que a hospeda (teoria da endosimbiose) por substâncias orgânicas como a glicose, as quais processa e converte em energia sob a forma de ATP, que devolve para a célula hospedeira, sendo energia química que pode ser usada em reações bioquímicas que necessitem de gasto de energia.
Formada por uma membrana dupla, que se organiza em cristas e uma região central, apresenta ribossomos soltos em seu interior, apresenta seu próprio DNA.
The circular 16,569 bp human mitochondrial genome
encoding 37 genes, i.e., 28 on the H-strand and 9 on the L-strand.
As mitocôndrias podem ter formatos variados. Uma mitocôndria contém membranas externas e internas compostas por bicamadas fosfolipídicas e proteínas. As duas membranas têm propriedades diferentes. Devido a esta organização de membrana dupla, existem cinco partes distintas para uma mitocôndria são elas:
Membrana mitocondrial externa,
Espaço intermembrana (o espaço entre as membranas externa e interna),
Membrana mitocondrial interna,
Espaço das cristas (formado por dobras da membrana interna), e
Imagem STEM (microscópio eletrônico de transmissão de varredura) de um cloroplasto. Figura superior: Fatia tomográfica de STEM de 10 nm de espessura de um cloroplasto de alface. As pilhas de Grana são interconectadas por tilacóides do estroma não empilhados, chamados de “lamelas do estroma”. Inclusões arredondadas associadas aos tilacóides são plastoglóbulos. Barra de escala = 200 nm. Figura inferior: Modelo 3D em grande escala gerado a partir da segmentação de reconstruções tomográficas por STEM. grana em amarelo; lamelas do estroma em verde; plastoglóbulos em púrpura; envelope de cloroplasto em azul.
(STEM (Scanning transmission electron microscope) imaging of a chloroplast
(Top) 10-nm-thick STEM tomographic slice of a lettuce chloroplast. Grana stacks are interconnected by unstacked stromal thylakoids, called “stroma lamellae”. Round inclusions associated with the thylakoids are plastoglobules. Scalebar = 200 nm. (Bottom) Large-scale 3D model generated from segmentation of tomographic reconstructions by STEM. grana = yellow; stroma lamellae = green; plastoglobules = purple; chloroplast envelope = blue).
Estrutura de montagem granum-estroma. O modelo predominante da montagem granum-estroma são pilhas de tilacóides granais envoltos por tilacóides estromais helicoidais destros que estão conectados a grandes folhas paralelas de tilacóides estromais e hélices destras adjacentes por estruturas helicoidais canhotas.
(Granum-stroma assembly structure The prevailing model of the granum-stroma assembly is stacks of granal thylakoids wrapped by right-handed helical stromal thylakoids which are connected to large parallel sheets of stromal thylakoids and adjacent right-handed helices by left-handed helical structures.)
As reaçõs da fase luminosa da fotossíntese reações luminosas da fotossíntese que acontecem no tilacoide (Fonte: Somepics)
Cloroplastos são organelas que realizam fotossíntese, onde a clorofila do pigmento fotossintético captura a energia da luz solar ou fóton, e armazena nas moléculas de armazenamento de energia ATP e NADPH enquanto libera o oxigênio da água para o meio.
Os cloropolastos são organelas que podem ser encontrdas em Protoctistas, como algas unicelulares e pluricelulares e plantas.
Esses compostos ricos em energia: ATP e o NADPH, são usados para sintetizar moléculas orgânicas a partir do dióxido de carbono em um processo conhecido como ciclo de Calvin.
O ciclo de Calvin-Benson. O ciclo de Calvin Benson é a fase não dependente da luz da fotossíntese, que ocorre no estroma do cloroplasto (liquido que envolve os tilacóides). E incorpora dióxido de carbono em moléculas de açúcar.
Nesta segunda etapa (fase química), é utilizado o ATP e o NADPH produzidos na etapa fotoquímica.
O dióxido de carbono combina-se com uma pentose (composto de 5 C) chamado de ribulose difosfato (RuDP), originando um composto intermédio, instável, com seis carbonos.
Devido à sua instabilidade, este composto origina de imediato duas moléculas de fosfoglicerato ou ácido fosfoglicérico (PGA), constituído por 3 carbonos.
Estas duas moléculas são fosforiladas pelo ATP e posteriormente reduzidas pelo NADPH, a partir da fase dependente da luz. Estas reacções formam o aldeído fosfoglicérico (PGAL). Por cada doze moléculas de PGAL formadas, dez são utilizadas para regenerar a ribulose (fazendo assim voltar, após a fosforilação deste por 6 ATPs, o ciclo à etapa inicial). Duas moléculas são utilizadas para formar compostos orgânicos, como glícidos e outros, dando assim sentido à autotrofia da fotossíntese. As moléculas de ácido pirúvico resultantes da degradação da glicose penetram no interior das mitocôndrias, onde ocorrerá a respiração propriamente dita. Cada ácido pirúvico reage com uma molécula da substância conhecida como coenzima A, originando três tipos de produtos: acetil-coenzima A, gás carbônico e hidrogênios.
Os cloroplastos realizam várias outras funções, incluindo a síntese de ácidos graxos, grande parte da síntese de aminoácidos e a resposta imune nas plantas. O número de cloroplastos por célula varia de um, em algas unicelulares, até 100 em plantas como Arabidopsis sp e Triticum sp.
Os cloroplasto é um tipo de organela conhecido como plastídio, caracterizado por suas duas membranas e uma alta concentração de clorofila. Outros tipos de plastídios, como o leucoplasto e o cromoplasto, contêm pouca clorofila e não realizam a fotossíntese.
Tipos de plastídios. As plantas contêm muitos tipos diferentes de plastídios em suas células.
Tabela da presença de vários pigmentos através dos grupos de cloroplastos. Células coloridas representam a presença
Delesseria sanguinea, uma especie de alga vermelha, que apresneta cloroplastos que contêm pigmentos vermelhos, como ficoerterina, que mascaram sua clorofila a azul-esverdeada. (Fonte: Gabriele Kothe-Heinrich).
(Delesseria sanguinea, a red alga, has chloroplasts that contain red pigments like phycoerytherin that mask their blue-green chlorophyll a)
A ficoeritrina (PE) é um complexo proteína-pigmento vermelho da família da ficobiliproteína coletor de luz, presente em algas vermelhas e criptófitas (grupo de algas com plastídeos Criptomonas sp). Esse pigmento é acessório aos principais pigmentos de clorofila responsáveis pela fotossíntese.
Como todas as ficobiliproteínas, é composto por uma parte da proteína ligada covalentemente à cromóforos chamadas ficobilinas. Na família da ficoeritrina, as ficobilinas mais conhecidas são: ficoeritrobilina, o cromóforo aceptor típico da ficoeritrina e, às vezes, a ficourobilina.
Ficoeritrinas são compostas por monômeros (αβ), geralmente organizados em um trímero em forma de disco (αβ) 3 ou hexâmero (αβ) 6 (o segundo é a unidade funcional das hastes da antena). Esses complexos típicos também contêm um terceiro tipo de subunidade, a cadeia γ. (Ficoeritrina)
Os cloroplastos são altamente dinâmicos, eles circulam e se movem dentro das células vegetais e, ocasionalmente, dividem-se em dois na reprodução.
Seu comportamento é fortemente influenciado por fatores ambientais como cor e intensidade da luz. Os cloroplastos, como as mitocôndrias, possuem seu próprio DNA, que se acredita ter sido herdado de seu ancestral, uma cianobactéria fotossintética que foi englobada por uma célula eucariótica primitiva.
Os cloroplastos não podem ser produzidos pela célula vegetal e devem ser herdados por cada célula filha durante a divisão celular.
Peroxissomos
Organelas especializadas formadas por uma só membrana, responsáveis por processos oxidativos da células. Peroxissomos são bolsas membranosas que contêm certos tipos de enzimas digestivas.
Sua semelhança com os lisossomos fez com que fossem confundidos com eles até bem pouco tempo. Entretanto, hoje se sabe que os peroxissomos diferem dos lisossomos principalmente quanto ao tipo de enzimas que possuem. Os peroxissomos, além de conterem enzimas que degradam gorduras e aminoácidos, contém também grandes quantidades da enzima catalase.
A catalase é a denominação dada para uma enzima fabricada por todos os seres vivos. Ela tem a responsabilidade de decompor o peróxido de hidrogênio e pode ser encontrada na organela conhecida como peroxissomo, que são encontrados nas células vivas.
A boa atividade do fígado e dos rins depende muito da função da catalase. Isso porque neles são encontrados peroxissomos em abundância, que são responsáveis por parte da desintoxicação do organismo.
A atuação da catalase e os peroxissomos contribuem para a fabricação dos sais biliares e também para a inutilização das substâncias tóxicas, no fígado.
A catalase é uma das enzimas mais eficientes encontradas nas células. Somente uma molécula dela é capaz de decompor milhões de moléculas de peróxido de hidrogênio. (Educa)
A catalase converte o peróxido de hidrogênio, popularmente conhecido como água oxigenada (H2O2), e água e gás oxigênio. A água oxigenada se forma normalmente durante a degradação de gorduras e de aminoácidos, mas, em grande quantidade, pode causar lesões à célula.
Apesar das descobertas recentes envolvendo os peroxissomos, a função dessas organelas no metabolismo celular ainda é pouco conhecida. Entre outras funções, acredita-se que participem dos processos de desintoxicação da célula.
Cloroplasto e um peroxissomo em uma célula vegetal
Citoesqueleto
O citoesqueleto é uma estrutura celular, em formato de rede no interior do citoplasma da célula, composta por um conjunto de três tipos diferentes de filamentos protéicos. São eles: microtúbulos, filamentos intermediários e microfilamentos. O citoesqueleto esta formado basicamente por duas proteínas estruturais: actina e tubulina.
Elementos dos citoesqueleto
Elementos do Citoesqueleto
Microtúbulos
São tubos ocos e compridos e são maiores do que os demais componentes do citoesqueleto. São eles os responsáveis tanto pela forma celular quanto pela movimentação que ocorre no interior das células. Agem também na movimentação dos cílios e flagelos.
Filamentos intermediários
Os filamentos intermediários estão no meio termo, em relação a sua forma, se comparados com os microtúbulos e microfilamentos. Sendo mais grossos do que os microfilamentos e mais fino do que os microtúbulos. Como formam uma rede internamente na célula tem a função de manter as organelas celulares em seus lugares.
Microfilamentos
os microfilamentos ajudam a manter o vigor e a forma celular, a turgescência; dão suporte mecânico e colaboram na movimentação; contribuem com funções da membrana plasmática; agem na contração muscular; migração de células embrionárias; combate a infecções e nos processos de cicatrização da pele.
Funções
Manutenção e organização celular interna e externa, tanto em sua forma quanto em seu conteúdo.
É responsável também pela movimentação das células;
Dá forma a célula;
Possibilita o movimento circular do citoplasma no interior da célula, atuando no processo de transporte de substâncias;
Permite a união das células, na formação de tecidos;
O citoesqueleto das células presentes nos músculos atua no processo de contração muscular;
Formação e movimentação de flagelos e cílios;
Em amebas e algumas espécies de protozoários, o citoesqueleto é responsável pela movimentação ameboide.
No processo de divisão celular eucariota, participam da movimentação dos cromossomos. (TodaBiologia)
Citoesqueleto e organização tecidual
Composição da ECM (matriz extra celular)
Tipos de colágenos do tecido conjuntivo
Colágeno I: 90% do total de colágeno do corpo. Principalmente na pele e osso
ColágenoII: essencilmente na cartilagem
Colágeno III: derme e paredes de vasos sanguíneos (ex. artéria aorta)
Colágeno IV: suporte e forma a membrana basal (formação dos tecidos), membrana basal testicular
Colágeno V: flexibilidade e resistência aos tecidos, encontrado nos ossos, tendoões, sangue e placenta.
Colágeno IX
Colágeno XI
Tipos de colágeno
Colágeno Tipo I
É o mais comum; aparece nos tendões, na cartilagem fibrosa, no tecido conjuntivo frouxo comum, no tecido conjuntivo denso (onde é predominante sobre os outros tipos), sempre formando fibras e feixes, ou seja, está presente nos ossos, tendões e pele. As espessas fibras de colágeno organizadas paralelamente são responsáveis pela grande resistência dos tendões.
Colágeno Tipo II
É produzido por condrócitos, aparece na cartilagem hialina e na cartilagem elástica. Não produz feixes. Presente nos discos intervertebrais, olhos e cartilagem. A suplementação de Colágeno tipo II é indicado para tratamento de artrose, por ajudar a recompor a cartilagem das articulações.
Colágeno Tipo III
Presentes em músculo liso, endoneuro e nas trabéculas dos órgãos hematopoieticos (baço, nódulos linfáticos, medula óssea vermelha); artérias, fígado, útero e camadas musculares do intestino.Abundante no tecido conjuntivo frouxo, é encontrado na artéria aorta do coração, nos pulmões, nos músculos dos intestinos, fígado, no útero. Constitui as fibras reticulares.
Colágeno Tipo IV
Aparece na lâmina basal, um dos componentes da membrana basal dos epitélios. Presente nas lentes da cápsula ocular, glomérulos.
Colágeno Tipo V
Está presente na maioria dos tecidos localizados no interior, especialmente na cobertura de vários elementos funcionais do corpo, tais como órgãos. Sua missão é dar a essas partes do corpo o poder de esticar e resistir a todos os tipos de fatores.
Colágeno Tipo VI
Está presente no sangue, camada íntima da placenta.
Colágeno Tipo VII
Está presente nas membranas corioaminióticas e na placenta.
Colágeno Tipo VIII
Está presente no endodélio..
Colágeno Tipo IX
Ele tem a função de manter as células unidas e é o principal componente proteico de órgãos
Colágeno Tipo X, XI e XII
Está presente na cartilagem.
Discussão
Um modelo atual muito difundido da evolução dos primeiros organismos vivos é que eles eram alguma forma de procariotos, que podem ter evoluído a partir de protocélulas, enquanto os eucariotos evoluíram posteriormente na história da vida.
Alguns autores questionam essa conclusão, argumentando que o conjunto atual de espécies procarióticas pode ter evoluído de ancestrais eucarióticos mais complexos por meio de um processo de simplificação. Outros argumentaram que os três domínios da vida (Bacteria, Archaea e Eukarya) surgiram simultaneamente, a partir de um conjunto de células variadas que formaram um único pool genético.
Essa controvérsia pode ser resumida da seguinte forma:
Não há consenso entre os biólogos sobre a posição dos eucariotos no esquema geral da evolução celular. As opiniões atuais sobre a origem e posição dos eucariotos abrangem um amplo espectro, incluindo as visões de que os eucariotos surgiram primeiro na evolução e que os procariotos descendem deles, que os eucariotos surgiram simultaneamente com eubactérias e arqueobactérias e, portanto, representam uma linha primária de descendência de igual idade e posição como os procariontes, que os eucariotos surgiram por meio de um evento simbiótico que implica uma origem endossimbiótica do núcleo, que os eucariotos surgiram sem endossimbiose e que os eucariotos surgiram por meio de um evento simbiótico que implica uma origem endossimbiótica simultânea do flagelo e do núcleo, além de muitos outros modelos.
Maneiras de passar através de uma menbrana plasmátaica
A membrana plasmática é de fundamental importância para a vida, uma vez que delimita o espaço interno da célula, isolando o meio intracelular do ambiente externo.
Esse isolamento, porém, não pode ser absoluto: para viver, a célula precisa permitir a entrada de certas substâncias úteis, como a água, gás oxigênio, gás carbônico, glicose, aminoácidos, alimento, etc.
A membrana plasmática desempenha uma função vital básica: através da permeabilidade seletiva permite a passagem de solutos (substâncias sólidas e líquidas).
Tramporte através da membrana
O transporte através da membrana pode ser dividido em transporte passivo e transporte ativo.
O transporte passivo divide-se por sua vez em difusão simples, difusão facilitada e osmose.
Já o transprote ativo pode ser primário ou secundário.
Transporte passivo é o transporte que ocorre entre uma membrana que separa duas soluções e tem por objetivo igualar as concentrações. Esse tipo de transporte ocorre sem gasto de energia e pdoe ser dividido em: difusão simples, difusão facilitada e osmose.
O transporte passivo é a passagem de uma substância, através da membrana, de uma região onde essa substância está mais concentrada para uma onde está menos concentrada, sem gasto de energia.
Existem três tipos de transporte passivo pela membrana celular: a difusão simples, a difusão facilitada e a osmose.
A difusão simples ocorre com o movimento de partículas através da membrana plasmática de um meio onde essas partiulas estão mais concentradas para um meio onde estão menos concentradas.
Condições
Para que a difusão possa efetivamente ocorrer essas substâncias devem ser pequenas o suficiente para poder atravessar a membrana.
A membrana deve ser permeável a elas.
É assim que acontece geralmente a difusão de gases, como o oxigênio e o gás carbônico através da membrana.
Difusão facilitada. Nesse processo, as substâncias podem difundir-se ainda de acordo com um gradiente de concentração, do meio mais concentrado para o menos concentrado, utilizando-se de proteínas especiais que atuam como canais, ou poros, permitindo a passagem das substâncias, denominadas de porinas. uma delas é a porina da água que é chamada de aquaporina.
Difusão simples
O soluto penetra na célula quando sua concentração é menor no interior celular do que no meio externo e sai da célula no caso contrário. Neste processo não há consumo de energia. Ocorre a favor do gradiente.
Algumas substâncias como a glicose, galactose e alguns aminoácidos são grandes em relação aos poros da membrana e não são solúveis em lipídios, o que também impede a sua difusão pela matriz lipídica da membrana. No entanto, essas substâncias passam através da matriz, por transporte passivo, contando, para isto, com o trabalho de proteínas carregadoras (proteínas transportadoras).
Osmose
Já a osmose é a passagem de solvente (água) de uma solução menos concentrada para uma solução mais concentrada, através de uma membrana semipermeável, até que a pressão exercida pela solução sobre a membrana impeça a passagem de solvente.
comportamento da célula em diferentes meios com difetentes concentrações de soluto
Na osmose ocorre a difusão (movimento) das moléculas de água (solvente) para o meio onde há maior concentração de solutos (sais, açúcares etc). Uma pressão atua equilibrando a entrada de água na célula dependendo da quantidade de soluto ali presente, denominada de pressão osmótica.
Num meio isotônico cuja concentração de solutos for igual a concentração desse soluto no citoplasma, ou seja, a concentração de solutos no meio intracelular e no meio extracelular são equivalentes, a pressão osmótica entre eles é igual, não havendo perda nem ganho de água entre eles.
No meio hipertônico, i.e., Quando há maior concentração de solutos no meio extracelular, esse meio exercerá maior pressão osmótica (hipertônico) sobre o meio intracelular (hipotônico) e a célula perderá água.
Num meio hipotônico i.e., quando há menor concentração de solutos no meio extracelhar, esse meio exercerá menor pressão osmótica (hipotônico) sobre o meio intracelular (hipertônico) e a célula ganhará água, podenso ocorrer a lise celular pela entrata em excesso de água.
Passagem da água (solvente) de um meio menos concentrado para o meio mais concentrado
Passagem da água (solvente) de um meio menos concentrado para o meio mais concentrado.
Transporte ativo
O transporte ativo é realizado por proteínas carreadoras e utiliza energia sob a forma de ATP para realizar o transporte de íons e outras substâncias através da membrana plasmática contra um gradiente de concentração, ou seja, é neste tipo de transporte que um íon localizado em uma região de baixa concentração pode ser transportado para outra com alta concentração deste mesmo íon, mantendo desta maneira uma manutenção na diferença de concentração de cátions e ânions de forma diferenciada no lado intra e extracelular.
O transporte ativo pode ser classificado em primário quando a proteína transportadora utiliza energia a partir de uma reação química exotérmica, e secundário quando o movimento independe diretamente do ATP e está associado à diferença de concentração de íons estabelecida pelo transporte ativo primário. (infoescola).
Endocitose
Endocitose é o processo pelo qual as células vivas ativamente absorvem material (moléculas, matéria orgânica, líquidos ou mesmo outras células) através da membrana plasmática.
Fagocitose, Pinocitose e Endocitose mediada por receptor
É a entrada de substâncias em uma célula por englobamento das particulas pela membrana celular.
Podemos distinguir tres tipos de endocitose
Fagocitose
A fagocitose é o processo pelo qual a célula, engloba partículas sólidas geralmente graças à formação de expansões da membrana denominadas pseudópodes. O material ingerido fica totalmente envolto pela membrana formando o fagossomo.
Pinocitose
Pinocitose, é processo pelo qual a célula, engloba partículas inespecíficas em meio aquoso, ou gotículas líquidas, por meio da invaginação na membrane plasmática. Quando a membrana se fecha sobre si forma-se uma vesícula chamada pinossomo que é puxada pelo citoesqueleto para o hialoplasma.
Endocitose mediada por um receptor
Endocitose mediada por um receptor, consiste na ligação de uma molécula extracelular a um receptor na membrana celular. Estes receptores, igualmente constituintes da membrana, estão muitas vezes associados à proteína do citoplasma denominada clatrina que forma uma depressão na membrana; quando um receptor se liga a uma molécula, a depressão aumenta até se transformar num vacúolo rodeado de clatrina, que entra na célula puxada pelo citoesqueleto.
Receptor-mediated endocytosis (RME), also called clathrin-mediated endocytosis
Esse processo forma vesículas contendo as substâncias absorvidas e é estritamente mediado por receptores na superfície da célula. Somente as substâncias específicas do receptor podem entrar na célula por meio desse processo.
A seletividade e o transporte através da membrana
A seletividade da membrana plasmática, representada pela capacidade da membrana em transportar determinadas substâncias, constitui uma das suas propriedades mais notáveis (Fig. abaixo). Pequenas moléculas ou íons passam pela membrana por simples difusão ou por meio de proteínas integrantes que mediam esse transporte. Estruturas maiores, como macromoléculas ou até organismos inteiros, como vírus e bactérias, entram na célula usando a rota endocítica, mediada por proteínas receptoras presentes na membrana.
Esquema que ilustra a seletividade de uma membrana sintética constituída apenas por uma bicamada lipídica. Algumas moléculas passam facilmente, enquanto que outras só passam através da mediação de proteínas transportadoras integrantes de membrana.
(Fonte: ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008).
O transporte mais simples que se pode conceber é a difusão simples, i.e., a difusão física das substâncias através da bicamada lipídica da membrana plasmática (Fig. abaixo). Também chamado de transporte não mediado, a difusão depende, basicamente, da existência de um gradiente de concentração favorável e da natureza da substância a ser transportada.
De um modo geral, quanto maior for sua solubilidade em lipídios e menor seu tamanho molecular, mais facilmente se dará sua passagem pela membrana.
A água constitui uma exceção notável: apesar da barreira hidrofóbica, suas moléculas difundem-se de forma relativamente rápida através da membrana.
A simples difusão de moléculas lipossolúveis ou de pequenas moléculas polares, mas não carregadas, como a água e o glicerol ocorre, portanto, sem grandes problemas (Fig. acima). No entanto, devido ao seu interior hidrofóbico, a membrana plasmática constitui uma séria barreira à passagem da maioria das moléculas maiores não carregadas, como a glicose e a sacarose, assim como íons.
Como o transporte de muitas dessas substâncias, que incluem também aminoácidos e nucleotídios, por exemplo, é vital para a célula, uma série de mecanismos foram desenvolvidos no sentido de permitir ou mesmo auxiliar sua passagem, especificamente, através dessa barreira.
Esquema dos principais tipos de transporte de substâncias através da membrana plasmática. (Fonte: COSTA, S.O.P. (coord.) - Genética Molecular e de Microorganismos. São Paulo, Manole, 1987).
Classificação dos tipos de transporte através da membrana plasmática
(modif. de Costa, S.O.P, 2008)
Contudo, a célula precisa de algumas moléculas que não são capazes de atravessar a membrana plasmática por simples difusão, como aminoácidos e nucleotídeos. Deste modo, é vital para a célula que ela possua uma série de mecanismos que permitam ou auxiliam sua passagem, especificamente, através dessa barreira (Fig. acima). Tais mecanismos envolvem determinadas proteínas integrantes de membrana, que mediam a passagem dessas substâncias. Algumas dessas proteínas, conhecidas por proteínas de canal, formam, como o próprio nome está dizendo, canais internos ladeados por aminoácidos hidrofílicos que permitem a passagem, por difusão, de certos solutos de tamanho apropriado.
Essas proteinas são formadas sobretudos por motivos do tipo beta-filha e barris beta, com os aminoácidos hidrofílicos situados na superfície interna do barril.
Alguns canais encontram-se permanentemente abertos (canais livres), pouco comuns na membrana plasmática. Na maioria dos casos, os canais se abrem após algum estímulo recebido pela célula.
Outras proteínas, denominadas de proteínas transportadoras ou permeases, atuam de modo bem mais sofisticado, executando um tipo de transporte denominado de transporte mediado.
Neste tipo de transporte ocorre um reconhecimento inicial do soluto a ser transportado (agindo, pois, como ligante), que se acopla ao sítio ativo da permease de maneira análoga à ligação enzima-substrato. Após esta ligação, a proteína transportadora sofre uma alteração conformacional, de tal forma que seu sítio ativo fica, agora, voltado para o lado oposto da membrana. Ocorre então o desacoplamento do soluto, completando-se sua transferência (Fig. acima num. 3).
O transporte mediado pode ser passivo, quando não houver gasto de energia metabólica para sua efetivação; neste caso, a transferência de soluto só se dará a favor de um gradiente de concentração.
O transporte mediado também poderá ser ativo, com gasto de energia metabólica e contra um gradiente de concentração. Um exemplo clássico desta última situação é proporcionado pela chamada "bomba de sódio-potásio" (bomba Na+- K+), encontrada em, virtualmente, todas as células animais (Fig. abaixo).
Esquema da “Bomba” (ATPase) de Na+ e K+. Gradiente eletroquímico de Na+; sítio de ligação do Na+; K+ sítio de ligação do K+; citossol; Gradiente eletroquímico de K+.
O esquema acima ressalta o funcionamento da “Bomba” (ATPase) de Na+ e K+, mostrando a atuação do gasto de ATP para colocar três moléculas de Na+ para fora da célula via um mecanismo de bombeamento contra o gradinte de concentração do sódio; e captura de duas moléculas de potásio que são trazidas para dentro da célula pela mesma bomba.
(Fonte: ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 4th Edition, New York, Garland, 2002).
Representada por uma proteína enzimática integrante de membrana: Na+-K+ATPase, agindo como permease, a "bomba" de Na+-K+ elimina ativamente íons Na+, enquanto, ao mesmo tempo, concentra íons K+ no interior da célula, com hidrólise simultânea de ATP. Tal sistema de transporte tem uma importância fundamental na geração e manutenção do potencial elétrico existente através da membrana, essencial para o desempenho de muitas funções celulares. (E-disciplinas).
Sabemos que a membrana plasmática é o limite semipermeável com permeabilidade seletiva, e é responsável,entre outras funções, por controlar todas as substâncias que entram e saem de uma célula.
A membrana plasmática é a principal interface celular, através da qual o interior da célula se comunica com o meio externo. O primeiro contato da célula com qualquer material extracelular é através de seus componentes lipídicos (ex. fosfolipídios) e proteicos (ex. receptores membranares).
Sendo assim, um dos primeiros testes para avaliar a interação dos nanomaterias com células eucarióticas é avaliar a integridade da membrana. Para isso é possível dosar a presença ou atividade de enzimas intracelulares (ex. lactato desidrogenase) ou testar a integridade da membrana com moléculas que não a permeiam (ex. azul de tripano ou iodeto de propídio).
Por exemplo, Carlson e colaboradores demonstraram que nanopartículas de prata (diâmetro de 15 a 55 nm) causaram danos à membrana, os autores avaliaram o extravasamento da enzima lactato desidrogenase para o meio extracelular, levando macrófagos à apoptose em menos de 24 h. Os autores sugeriram que a perda da integridade de membrana foi responsável pela morte celular.
Em virtude dessa capacidade, (controle de entrada e saída) dizemos que a membrana apresenta permeabilidade seletiva. Essa seleção é importante porque garante a entrada de oxigênio e nutrientes na célula, além de garantir a eliminação dos produtos do metabolismo celular.
TIPOS DE TRANSPORTES ATRAVÉS DA MEMBRANA
O transporte passivo é a passagem de uma substância através da membrana plasmática de uma região onde ela está mais concentrada para uma onde está menos concentrada, sem gasto de energia. Existem três tipos de transporte passivo pela membrana celular: a difusão simples, a difusão facilitada e a osmose.
Difusão simples
A difusão simples ocorre com o movimento de partículas de um meio onde estão mais concentradas para um onde estão menos concentradas. No entanto, essas substâncias devem ser pequenas o suficiente para atravessar a membrana, e a membrana deve ser permeável a elas. É assim que acontece geralmente a difusão de gases, como o oxigênio e o gás carbônico.
Do meio mais concentrado para o menos concentrado
Nesse processo, as substâncias podem difundir-se ainda de acordo com um gradiente de concentração, do meio mais concentrado para o menos concentrado utilizando-se de proteínas que atuam como canais, ou poros, permitindo a passagem das substâncias, essas proteinas são denominadas de porinas. (A)
Um barril beta é uma folha beta composta de repetições em tandem que se torce e se enrola para formar uma estrutura toroidal fechada na qual a primeira fita é ligada à última fita (ligação por pontes de hidrogênio).
As fitas beta em muitos barris beta são organizadas de forma antiparalela.
As estruturas do barril beta são assim chamadas pela sua semelhança com os barris usados para conter líquidos.
A maioria delas são proteínas solúveis em água e freqüentemente se ligam a ligantes hidrofóbicos no centro do barril, como nas proteínas lipocalinas (familia de proteinas as quais transportam pequenas moléculas hidrofóbicas tais como esteróides, retinóis, e lipídeos).
Outros se atravessam membranas celulares e são comumente encontrados nas porinas. Estruturas de barril semelhantes a porinas são codificadas por até 2–3% dos genes em bactérias Gram-negativas.
Barril β de 18 fitas.
Uma Porina específica para sacarose bacteriana de Salmonela typhimurium.
Este barril β fica em na membrana e permite que a sacarose se difunda do meio extracelular para o citoplasma. (Fonte: Por Opabinia regalis)
Retinol-binding protein in a calculated membrane-bound state of the protein 1kt6
Na difusão facilitada uma substância é transportada por meio da participação de proteínas presentes na membrana. Essas proteínas são conhecidas como proteínas carregadoras e facilitam o movimento espontâneo das moléculas sem que haja nenhum gasto de energia pela célula. Substâncias como aminoácidos e açúcares podem ser transportadas dessa forma para o interior da célula.
Na difusão facilitada, proteínas reconhecem a substância a ser transportada em um mecanismo de “chave e fechadura”, permitindo a entrada dessa substância na célula.
Osmose
A osmose é um tipo especial de difusão e, nesse caso, a substância que se difunde pela célula é a água. Nas células, a água difunde-se do meio menos concentrado para o mais concentrado. Se uma célula é colocada, por exemplo, em um meio em que a concentração de soluto (substância dissolvida) é muito maior que a do interior da célula, a tendência da célula é perder água por osmose. Se o contrário ocorre, a célula enche-se de água. (B)
Difusão da água do meio menos concentrado para o mais concentrado
A osmose é o movimento de água através de uma membrana semipermeável no caso a membrana plasmática da célula ocasionado por diferenças na pressão osmótica.
É uma característica muito importante para a manutenção da vida das células.
As membranas plasmáticas são mais permeáveis à água que a maioria das outras moléculas pequenas, íons e macromoléculas, por que os canais protéicos, as aquaporinas presentes na membrana, seletivamente permitem a passagem de água. Na região interna da aquaporina estão presentes aminoácidos hidrofílicos, que impulsionam a água do meio menos concentrado para o meio citoplasmático, mais concentrado. No caso dos protozoários, eles não tem como evitar a entrada da água, assim a seleção dotou-os de um vacúolo contrátil ou pulsátil que joga para fora a água em excesso.
Nesse caso (osmose) a água move-se, sem nenhum gasto de energia para dentro da célula, indo do do meio menos concentrado para o mais concentrado, o citoplasma, através da membrana plasmática que é seletivamente permeável.
Se quisermos interromper a osmose, fluxo natural da água do meio menos concentrado para o mais concentrado, é necessário exercer sobre o sistema formado por duas soluções ou uma solução e um solvente, separados por uma membrana semipermeável, uma pressão no sentido inverso ao da osmose no mínimo com a mesma intensidade daquela que o solvente faz para atravessar a membrana semipermeável.
A essa pressão, capaz de impedir o fenômeno da osmose, chamamos de pressão osmótica.
E pode ser definida como o equivalente à pressão necessária, aplicada sobre um recipiente contendo solvente puro de modo a impedir a osmose.
Osmose natural
É a saída do meio menos concentrado para o meio mais concentrado.
Pressão osmótica de uma solução
π = N.R.T.i
π = pressão osmótica da colução
N = Molaridade (concentração do soluto em solução expressa em mol/L)
R = constante universal dos gases perfeitos, cujo valores são 0,082 atm.L.K-1.mol-1 ou 62,3 mmHg.L.K-1.mol-1 ou 8,31 J/mol.K
T = temperatura em ºK
i= fator de correção de Van't Hoff
Solutos moleculares como a glicose e sacarose, de mesma concentração em mol/L, apresentam a mesma pressão osmótica.
Solutos iônicos como NaCl ou CaCl2, embora tenham a mesma concentração em mol/L, apresentam pressão osmótica diferentes, devido ao número de partículas produzidas. Como o NaCl se dissolve em água, gerando íons Na+ e Cl-, 1 mol/L de NaCl produz na verdade 2 mol/L de partículas.
No cálculo da Pressão Osmótica, o fator de Van't Hoff (i) adquire então valor 2. No caso do CaCl2, como temos 3 mol/L de partículas (Ca+ e 2Cl-), temos i = 3.
A medição da pressão osmótica pode ser usada para a determinação dos pesos moleculares.
A pressão osmótica é um fator importante que afeta as células.
Osmorregulação é o mecanismo de homeostase de um organismo para atingir o equilíbrio na pressão osmótica.
A hipertonicidade é a presença de uma solução que causa o perda de líquido das células.
A hipotonicidade é a presença de uma solução que causa o inchaço das células.
A isotonicidade é a presença de uma solução que não produz alteração no volume celular.
Quando uma célula está em um ambiente hipotônico, o interior da célula acumula água, a água flui através da membrana celular para dentro da célula, fazendo com que ela se expanda.
Nas células vegetais, a parede celular restringe a expansão, resultando em pressão de dentro da parede celular chamada de pressão de turgor, ou turgescência celular.
A pressão de turgor permite que as plantas herbáceas fiquem de pé. É também o fator determinante de como as plantas regulam a abertura de seus estômatos, por onde a água sai na forma de vapor.
Em células animais, a pressão osmótica excessiva pode resultar em citólise.
A pressão osmótica é a base da filtragem ("osmose reversa"), um processo comumente usado na purificação de água.
A água a ser purificada é colocada em uma câmara e submetida a uma pressão maior do que a pressão osmótica exercida pela água e pelos solutos nela dissolvidos.
Parte da câmara se abre para uma membrana diferencialmente permeável que permite a passagem das moléculas de água, mas não das partículas de soluto.
A pressão osmótica da água do oceano é de cerca de 27 atm.
A osmose reversa dessaliniza a agua do oceano produzindo água doce a paritr da agua salgada. Desta forma sendo uma maneira de produzir água doce a partir da água do oceano.
1. (UCPel) O esquema abaixo representa os tipos de transporte pela membrana.
Podemos afirmar que no tipo de transporte indicado pelo número:
a) 2, moléculas e íons tendem a seguir o seu próprio gradiente e ir em direção à região onde estão menos concentrados. A intensidade do gradiente, temperatura, tamanho do soluto, carga e pressão influenciam nessa taxa de transporte.
b) 1, os transportadores passivos permitem que íons ou moléculas pequenas atravessem uma membrana até o lado onde são menos concentrados. Como exemplo temos a bomba de sódio e potássio.
c) 1, uma proteína bombeia um soluto ao longo de uma membrana, contra seu gradiente de concentração. O transportador deve ser ativado, normalmente por um aporte de energia do ATP. Como exemplo temos transportador de glicose.
d) 2, um soluto se vincula a uma proteína que o libera no lado oposto da membrana. Nenhuma energia é necessária; o movimento líquido de soluto é para abaixo de seu gradiente de concentração.
e) 2, os transportadores ativos bombeiam íons ou moléculas através de membranas para o lado onde são mais concentrados. Exigem entrada de energia como do ATP.
2. (UEPG) A figura abaixo trata-se de uma representação esquemática da passagem de partículas de soluto através da membrana plasmática. Sobre o assunto, assinale o que for correto.
01. O transporte ativo, representado em D, ocorre através da membrana plasmática, com “gasto” de energia, ou seja, ocorre contra o gradiente de concentração.
02. Em A, B e C podemos observar exemplos de transporte pela membrana plasmática sem “gasto” de energia, tendendo a igualar a concentração da célula com a do meio externo, ou seja, acontece a favor do gradiente de concentração.
04. Em C, está representada a difusão facilitada. Neste processo, algumas proteínas da membrana, ou permeases, atuam facilitando a passagem de certas substâncias. Podemos citar, como exemplo, o transporte de glicose em células do fígado.
08. A difusão facilitada corresponde ao movimento de partículas de onde elas estão menos concentradas para uma região de maior concentração, como demostrado por D. Neste tipo de transporte, faz-se uso de energia para passagem de substâncias do meio intracelular (hipotônico) para o meio extracelular (hipertônico).
16. Na osmose, representada em A, partículas, íons e proteínas podem atravessar a membrana por simples difusão, com o objetivo da manutenção das concentrações em equilíbrio entre os meios intra e extracelular.
Os organismos vivos mantêm-se pela aquisição de nutrientes a paritr de seus ambientes. Os nutrientes abastecem com energia química e com os componentes moleculares para construir e manter um sistema vivo. Chamamos esses processos químicos essenciais de metabolismo.
Esses processos incluem digestão, a obtenção de energia (respiração) e síntese de moléculas e estruturas (células). O metabolismo é uma interação de reações destrutivas ou degradativas (catabólicas) com reações construtivas (anabólicas) ou de biossíntese.
Os processos químicos anabólicos e catabólicos mais fundamentais empregados pelos sistemas vivos apareceram cedo na história evolutiva da vida e são compartilhados por todas as formas vivas, i.e., são conservados ao longo da historia da vida em nosso planeta, desde as bactérias ao homem. Essas reações incluem a síntese de carboidratos, lipídios, ácidos nucleicos, proteínas e suas partes constituintes e a clivagem das ligações químicas, para recuperar a energia nelas estocada.
Nos animais, muitas reações metabólicas fundamentais ocorrem no nível celular,
frequentemente em organelas específicas, encontradas em todo o reino animal.
A respiração celular ocorre, por exemplo, nas mitocôndrias. As membranas celulares e nucleares regulam o metabolismo, controlando o movimento de moléculas através dos limites celular e nuclear, respectivamente. O estudo das funções metabólicas, desde os níveis bioquímicos (moleculares) aos do organismo, é conhecido como fisiologia. (1 para quem quiser baixar o livro grátis).
Hickman Jr, C.P. et alii. Princípios Integrados de Biologia. Rio de Janeiro, RJ, EDITORA GUANABARA KOOGAN LTDA, 2016
ANABOLISMO
O anabolismo é o conjunto de reações envolvidas na síntese de moléculas complexas, a partir de moléculas simples. Esse processo requer energia, e pode ser estimulado pela prática de exercícios e por uma alimentação rica em proteínas.
A construção de tecidos e o ganho de massa muscular são dois processos anabólicos básicos. Já no nível celular, um exemplo de processo anabólico é a sintetização de proteínas por meio de aminoácidos. (Diferença.com)
Alguns dos hormônios envolvidos no processo de anabolismo são:
Insulina, Estrogênio, Testosterona, Hormônio de crescimento, Esteroides.
O catabolismo é o conjunto de reações envolvidas na quebra de moléculas complexas em moléculas simples.
Essas reações liberam energia para o organismo, que posteriormente poderão ser usadas no processo anabólico. Alguns resíduos celulares como ureia, dióxido de carbono e amônia também são produzidos durante o catabolismo.
O principal processo catabólico é a digestão, onde as substâncias ingeridas pelo corpo são divididas em componentes mais simples. No nível celular, podemos citar a quebra de proteínas para a formação de aminoácidos, ou de glicogênio para a formação de glicose.
Alguns dos hormônios envolvidos no processo catabólico são:
Adrenalina, Cortisol, Glucagon, Citocinas.
D1BIO062 – Interpretar a equação química da respiração celular aeróbia.
A respiração aeróbica é representada pela seguinte equação:
C6H12O6 + 6 O2 + 6 CO2 + 6 H2O + Energia
Nessa equação é possível identificar que a molécula de glicose (C6H12O6) é dividida, de modo que possa originar as substâncias mais simples (CO2 e H2O).
Molécula de glicose e suas conformações: de Fischer e Haworth
Todas as reações da respiração celular ocorre dentro da mitocôndria, organela citoplasmática que atua como uma usina de energia da célula. Com isso, várias reações com a participação de enzimas e coenzimas nativas da mitocôndria são usadas para catalisar reações no processo que chamamos de respiração aeróbica. Essas reações favorece as constantes oxidações de glicose. Assim, esse modo de obtenção de energia gera trinta e seis (38) mols de ATP a partir de um mol de glicose. A respiração aeróbica se divide em três etapas: glicólise (ocorre no citoplasma), o Ciclo de Krebs e a fosforilação oxidativa (cadeia respiratória) ambos ocorrendo na mitocôndria.
D1BIO066 – Identificar as equações químicas gerais referentes à fermentação.
D1BIO067 – Distinguir fermentação lática e alcoólica.
O que é fermentação?
A produção da cerveja e vinho inicialemnte e depois de todas as outras bebidas alcoolicas, dependeu do desenvolvimento da agricultura e do cultivo de parreiras para uva e cereais como o trigo e a cevada.
A partir de aproximadamente 9 - 8 mil anos atrás o hábito de produzir e beber cerveja se espalhou, popularizando-se por todo o mundo.
Uma importante parte do processo cervejeiro não era compreendida e sua ocorrência era atribuída à mágica ou simplesmente ao divino, essa etapa era a fermentação. Os gregos deram o nome de Baco ao deus do vinho e da fermentação. No entanto esses povos ancestrais sabiam que tinham que executar as etapas do processo exatamente da mesma forma sempre para que a "magica" acontecesse no final: a produção da bebida. Uma vez amassada as uvas ou moida a cevada e misturada a água um processo desconhecido iniciava nos barris ou vasos que continam a mistura era a fermentação.
Na fermentação, a glicose é degradada parcialmente, na ausência de oxigênio, em substâncias orgânicas mais simples, como o ácido lático (fermentação lática) e o álcool etílico (fermentação alcoólica).
Glicose
Os monossacarídeos são os compostos mais simples de carboidrato, possuindo entre três e sete carbonos. São monômeros, ou seja, não podem ser hidrolisados. Geralmente possuem gosto doce e são sempre solúveis em água. Os mais conhecidos popularmente são as pentoses (5 carbonos) e hexoses (6 carbonos). São compostos de reserva das sementes ou dos frutos.
Os monossacarídeos apresentam a estrutura de um aldeído ou cetona que apresenta pelo menos dois grupos hidróxi, os açúcares que possuem carbonila de aldeído são chamados de aldoses, enquanto aqueles que possuem função cetona são chamados de cetoses. Podem ser nomeados de acordo com o comprimento da cadeia, 3, 4, 5 carbonos: triose, tetrose, pentose respectivamente.
A glicose é uma aldose, pois, conforme mostra a sua estrutura química, além dos grupos poliálcoois, ela possui um grupo aldeído em sua fórmula. Mais especificamente ela é uma aldo-hexose.
FERMENTAÇÃO
A fermentação, é um processo anaeróbico de síntese de ATP que não envolve a cadeia respiratória e tem como aceptor final de hidrogênios um composto orgânico o piruvato.
A fermentação é outra via anaeróbica (não requer oxigênio) para a quebra da glicose, uma via que é realizada por muitos tipos de organismos e células. Na fermentação, a única via de extração de energia é a glicólise, com uma ou duas reações extras acrescentadas ao final.
A fermentação e a respiração celular começam da mesma maneira, com a glicólise. No entanto, na fermentação, o piruvato produzido na glicólise não continua através da oxidação e ciclo do ácido cítrico, e acadeia transportadora de elétrons não acontece. Uma vez que a cadeia transportadora de elétrons não esta funcional, o NADH produzido na glicólise não pode entregar seus eletrons para voltar a forma NAS+. O propósito das reações extras na fermentação é regenerar o carreador de elétrons NAD+ a paritir do NADH produzido na glicólise.
As reações extras consegurm isso permintindo que o NADH entregue seus eletrons a uma molécula orgânica (o piruvato, o produto funal da glicólise). Isso permite qeu a glicólise permaneça funcionando pela garantia de um suprimento constante de NAD+.
Na fermentação do ácido lático, o NADH transfere seus elétrons diretamente ao piruvato, gerando o lactato como subproduto. O lactato, que é apenas a forma desprotonada do ácido lático, dá nome ao processo. As bactérias que produzem iogurte realizam fermentação do ácido lático, assim como as hemácias em nossos organismos, já que elas não possuem mitocôndrias, e portanto não podem realizar a respiração aeróbica. (11)
Ela ocorre no citosol da célula e é realizada por seres anaeróbicos, mas também pode ser uma alternativa de energia para os organismos aeróbicos em situações nas quais o gás oxigênio está ausente ou em níveis reduzidos (hipóxia).
Os organismos ancestrais se originaram em um mundo cuja atmosfera primitiva era redutora pois carecia de O2 e, por isso, pensa-se que a glicólise seja o mecanismo biológico mais primitivo para a obtenção de energia a partir de macromoléculas presentes em todas as formas de vida.
No curso da evolução, a química dessa sequência de reações foi completamente conservada. As enzimas glicolíticas dos vertebrados são muito semelhantes, tanto na sequência de aminoácidos quanto na estrutura tridimensional, àquelas das leveduras.
Enximas das leveduras semelhantes na sequência e estrutra às enzimas dos vertebrados.
A glicólise difere entre essas espécies apenas em detalhes de sua regulação e no destino metabólico do piruvato formado.
Se voce quer aprender mais sobre Fermentação e panificação:
Inicialmente acontece a glicólise, quando a molécula de glicose é degradada em dois piruvatos, cada um com três carbonos, com saldo de dois ATP.
Essa etapa é comum tanto para a fermentação como para a respiração.
O ácido pirúvico é convertido em etanol e CO2 em duas etapas
Fermentação alcoólica
1ª etapa: O ácido pirúvico é descarboxilado e forma-se acetaldeído;
2ª etapa: O acetaldeído é reduzido pelo NADH a etanol.
Os processos fermentativos envolvem conjuntos de reações enzimáticas que ocorrem no hialoplasma:
1)Glicólise:degradação da glicose em ácido pirúvico.
Na glicólise a glicose e é oxidada e formam-se duas moléculas de ácido pirúvico. O agente oxidante é o NAD+ que é transformado em NADH. O saldo energético é de duas moléculas de ATP.
2) Redução do ácido pirúvico
Conduz à formação dos produtos de fermentação.
O ácido pirúvico, ou moléculas orgânicas que se formam a partir dele, são aceptores dos elétrons do NADH, o que permite regenerar o NAD+.
O NAD+ pode, assim, voltar a ser utilizado na oxidação da glicose com formação de 2 ATP.
Os produtos finais da fermentação dependem da molécula orgânica que é produzida a partir do ácido pirúvico.
FERMENTAÇÃO ALCOÓLICA
COMPARANDO FERMENTAÇÃO COM RESIPIRAÇÃO
FERMENTAÇÃO LÁTICA
Na fermentação lática, o piruvato é transformado em lactato (ou ácido lático).
Os NADH formados na glicólise são reoxidados, perdendo elétrons e originando NAD+. Essa perda de elétrons fornece energia para a transformação do piruvato em lactato. O NAD+ é o aceptor final dos íons hidrogênios.
Desta forma, a equação simplificada da fermentação lática pode ser representada da seguinte maneira:
Algumas bactérias, protozoários e fungos realizam a fermentação lática. Mas o exemplo mais comum que temos são as nossas células musculares. A atividade física demanda alto consumo de energia, e a respiração celular é quem produz toda a energia necessária para sua realização.(Oshio, 2021,12)
A levedura Saccharomyces cerevisiae pode obter energia na ausência de oxigênio, de acordo com a equação
C6H12O6 → 2 CO2 + 2 CH3CH2OH + 2 ATP
Produtos desse processo são utilizados na indústria de alimentos e bebidas. Esse processo ocorre _____________ da levedura e seus produtos são utilizados na produção de _____________.
As lacunas dessa frase devem ser preenchidas por:
a) nas mitocôndrias; cerveja e vinagre. b) nas mitocôndrias; cerveja e pão. c) no citosol; cerveja e pão. d) no citosol; iogurte e vinagre. e) no citosol e nas mitocôndrias; cerveja e iogurte.
Comentários
Olha só que questão molezinha! O enunciado já começa falando sobre a levedura Saccharomyces cerevisae, aquele fungo que é utilizado no processo de fermentação de bebidas alcoólicas e na panificação. Se estamos falando de fermentação, o processo de obtenção de energia só pode ser anaeróbico.
Os organismos heterotróficos obtêm energia a partir da degradação de moléculas orgânicas obtidas do meio, em um processo chamado glicólise. A glicólise acontece no interior celular, no citosol. Lembre-se do quadro a seguir, pois ele é muito importante para “matar” várias questões:
Agora, vamos analisar as alternativas:
A alternativa A está errada, porque a fermentação ocorre no citosol das células. No caso dos organismos anaeróbicos facultativos (como a S. cerevisae), caso houvesse oxigênio no meio eles prosseguiriam para as demais etapas da respiração celular (que ocorrem na mitocôndria). Na ausência deste elemento, eles prosseguem transformando o produto da glicólise (piruvato) em etanol, realizando a fermentação alcoólica. Ainda, a fermentação alcoólica é o processo utilizado por essa levedura na fermentação de pães e bebidas. Já a produção de vinagre se dá por outro processo fermentativo, chamado fermentação acética.
A alternativa B está errada, porque a fermentação ocorre no citosol das células e não nas mitocôndrias.
A alternativa C está certa e é o nosso gabarito. O etanol produzido na fermentação da cevada está presente na cerveja e o dióxido de carbono liberado na fermentação promove o crescimento do pão.
A alternativa D está errada, porque a fermentação alcoólica é utilizada para a fermentação de bebidas alcoólicas, pães e como combustível. A fermentação lática é que é responsável pela produção de iogurte e outros produtos lácteos. A fermentação acética, como já foi dito, corresponde à transformação do álcool em ácido, conferindo o gosto característico de vinagre.
A alternativa E está errada, porque a fermentação não ocorre nas mitocôndrias e a fermentação alcoólica não produz iogurte.
Quando fazemos musculação e nossos músculos não respondem da maneira que deveriam, i.e., aumentando seu volume e força, pode estar ocorrendo o chamamos de catabolismo muscular.
Não é incomum que isso aconteça. Cada um de nós conhece ou tem um amigo que treina todo dia e não percebe alteração no "shape"; isso pode ocorrer por algumas razões entre elas: estar fazendo o exercício errado, exercícios sem a devida orientação, tempo excessivo, etc., o que acarreta no catabolismo.
O que ocorre é que o catabolismo atrofia os músculos e tecidos do corpo, muitas vezes por exagerar nos exercícios da academia, por má alimentação ou por falta de descanso adequado e até mesmo por alguma doença.
Por exemplo, uma virose, sobrecarrega os músculos trabalhados e não há tempo do organismo repor as energias necessárias para o funcionamento normal.
Toda pessoa que busca definição e hipertrofia muscular deve saber a dicotomia: anabolismo e catabolismo muscular.
O metabolismo tem dois momentos bem distintos: o anabolismo e o catabolismo. O anabolismo, conhecido como metabolismo construtivo contribui para o crescimento, regeneração e manutenção de tecidos e órgãos, entre eles os músculos. Isso porque o anabolismo é o conjunto de reações que colaboram para a construção das macromoléculas, i.e., para a biossíntese a partir de outras moléculas, resultando a manutenção dos órgãos. Portanto o anabolismo é a construção de tecidos entre eles os tecidos musculares. O catabolismo normal acontece a todo momento e é necessário para prover o organismo com a matéria prima para o anabolismo. O catabolismo muscular só acontecerá por motivos específicos como um perigo de fome muito grande, excesso de trabalho muscular, e grande estresse tecidual.
Para que não ocorra o catabolismo muscular é importante a ingestão de alimentos energéticos, como por exemplo alimentos ricos em polissacarídeos, polipeptídios, e lipídios.
Assim, a síntese de proteínas através de aminoácidos importantes para os músculos não estagnará, havendo construção e reparo de novo tecido muscular.
Portanto, o anabolismo acontece através da ingestão de nutrientes antes e pós-treino. Por isso, a alimentação é fundamental para a construção de músculos saudáveis, logo após um treino intenso é indispensável uma alimentação com carboidratos que serão convertidos em glicose e com proteínas que fornecerão os aminoácidos necessários para a hipertrofia muscular. (Blogeducaçãofísica)
Fatores que favorecem o catabolismo
1) Alimentação inapropriada: pobre e sem qualidade, treinamento longo e intenso em jejum, com fome, treino depois de duas horas após a última refeição ou uma alimentação sem carboidratos saudáveis pode acarretar em catabolismo. Assim como não consumir carboidratos e proteínas suficientes e de qualidade ou não se alimentar logo após-treino também resulta em catabolismo.
2) Treinar por longos períodos, ou seja, mais de 60 minutos e com treinos exaustivos e pesados também implica em catabolismo.
3) Descanso insuficiente: dormir pouco, ou não ter um período de descanso do músculo trabalhado facilita o catabolismo.
4) Ingestão de bebida alcoólica: interfere na capacidade do organismo em absorver nutrientes. Quando ingerimos bebida alcoólica, o nosso fígado passa trabalhar dobrado para eliminar as toxinas, e metabolizar o alcool. Para quem faz o consumo frequente de bebida alcoólica, estima-se uma perda de 20% de massa muscular e um aumento de gordura corporal.
Fatores que favorecem o anabolismo
Uma das principais contribuições para o anabolismo é a alimentação adequada e equilibrada, que forneça todos os nutrientes qeu nosso corpo necessita.
É recomendável que a proteína seja tanto anumal quanto vegetal, lembre-se dos nove (9) amonoácidos essenciais, que nosso corpo não produz e portanto ncessitamos obter da dieta, e dos 11 amnoácidos naturais ou não essenciais que nosso corpo produz. A esses alimentos devemos agregar carboidratos de preferência após o treinamento. A noite consuma alimentos preferencialmente sem o carboidrato, para evitar que nosso corpo acumule na forma de ácidos graxos.
Os principais alimentos anabólicos
Fontes de proteínas animais
Peito de frango
O peito de frango é uma das maiores fontes de proteína, em cada 100 gramas de peito de frango existem cerca de 32g de proteínas.
Outas carnes fontes de proteínas
Outras fontes de proteínas animais: alcatra bovina sem gordura grelhada 31,9g de proteina, costela suina 30,2g de proteína. Patinho sem gordura e grelhado 35,9 g de proteínas; lombo suíno assado 35,7 g de proteínas; contrafilé bovino sem gordura e grelhado 35,1 g de proteínas; sardinha assada 32,2 g de proteínas; fígado bovino grelhado 29,9 g de proteínas.
Carne vermelha
A carne vermelha bem como alimentos ricos em zinco como ostras, fígado, feijão, castanha ou sementes de girassol, contribuem para a produção natural da testosterona no corpo. Para obconseguir pruzir mais testosterona acrescente em sua dieta manda, tomate espinafre e óleo de peixe.
Alimentos ricos em zinco, vitamina A, vit. D e fosfolipídeos
Ovo
O ovo é um dos alimentos mais completos pois é rico em aminoácidos e proteínas alem de fosfolipídeos. Alem disso é um alimento com grande conteúdo de vitamina D. Alguns sintomas que podem indicar níveis de testosterona baixos incluem: pouco interesse sexual, esquecimento constante, cansaço frequente, alterações de humor, prevalecendo sintomas de depressão, Dificuldade em ter um corpo musculado, acúmulo de gordura, dificuldade em adormecer e muita agitação durante a noite, ossos frágeis.
Aveia
A avia fornece energia, saciedade e reduz a probabilidade de fadiga muscular.
Salmão
em 170 gramas do peixe contem cerca de 35% de proteínas, além de ácidos graxos e Omega 3.
Iogurte
o cálcio encontrado no leite controla a contração muscular e pode fazer com que o corpo se torne menos eficiente em armazenar gordura corporal. O cálcio possui a importante função de expor um sítio de ligação da miosina na proteína actina. Desta forma, o cálcio é fundamental para que ocorram as contrações musculares, sem este cátion não ocorrerá contração muscular.
Azeite de oliva extravirgem
O azeite de oliva tão caro à dieta mediterrânea, pode controlar inflamações no corpo. O azeite de oliva é rico em ácidos graxos monoinsaturados (ômega 9), o consumo regular de azeite contribui para a redução do colesterol ruim (LDL) e elevação do bom (HDL). Ele também é rico em polifenóis, compostos que respondem pelo sabor característico, além de terem ação antioxidante e preventiva de doenças cardiovasculares.2Níveis baixos de inflamação significam um aumento na recuperação muscular.
Axeite de oliva extra virgem
Outro fator importante para o anabolismo é um treino consciente e correto. Por isso, é importante focar na intensidade do exercício e não exagerar not empo. Isso significa que um treino de menos de 60 minutos será ideal quando vc chega no seu limite de peso e esforço. Todavia, depois da pandemia um treino mais longo com menos intensidade ajudará a perder os quilos extras adquiridos ao longo do tempo parado, longe da academia. Não podemos esquecer do descanso que é um fator importante para o bem estar dos músculos. Deve-se dormir pelo menos 8 horas, pois é durante o sono que os músculos crescem.
Descanso após o treino. O ideial é de 8 horas de sono diárias.
Pesquisa desenvolvida na Faculdade de Engenharia de Alimentos (FEA) da UNICAMP revela que três tipos de cerveja produzidos no país são uma fonte de vitamina B9, graças à presença dos folatos, compostos equivalentes ao ácido fólico. O estudo aponta que o consumo de 350 ml da bebida por dia poderia trazer benefícios à saúde humana, no combate às anemias, doenças cardiovasculares e malformações fetais, com aporte de cerca de 20% de folatos para o organismo.
A Agência Nacional de Vigilância Sanitária (Anvisa) recomenda o consumo de 400 microgramas de folatos por dia, também presente em outros alimentos. (FEA/UNICAMP)
Catabolismo muscular, por quê?
O catabolismo ocorre quando o corpo não encontra os nutrientes necessários na dieta diária ele então inicia a usar outras fontes de energia do nosso próprio corpo, ou seja, quando ficamos muito tempo sem nos alimentar, o nosso organismo que precisa de energia para atividades vegetativas de sobrevivência irá realocar essa energia que esta estocada primeiro na gordura (ácidos graxos e lipideos circulantes) depois na gordura estocada na forma de tecido adiposo, e depois nos nossos músculos. Assim, devemos estar atentos para não deixar o organismo sem energia por muito tempo fazendo-o entrar em uma rota catabólica, de degradação de tecidos.
Algumas dicas para evitar o catabolismo muscular
Fazer menos de 60 minutos de exercícios resistido intensos por dia.
Evite bebida alcoólica destilada e cerveja em excesso.
Evite o estresse que pode contribuir para acelerar o catabolismo muscular.
Doenças e viroses, podem contribuir para o catabolismo.
Poucas horas de sono. (Insônia desencadeia o catabolismo, busque tratamento, e tente melhora sua qualidade de vida; o sono é fundamental para os músculos se recuperarem. Dormir de 6 a 8 horas por dia é o ideal.
Evitar excesso de trabalho que é outro fator para reações catabólicas.
Evitar de ficar muito tempo sem se alimentar;
Alimentar-se corretamente antes e logo depois do treino.
Diminuir o número de repetições durante o treino resistido e compense com mais carga (e um menor número de repetições).
Ingerir muita proteína e também carboidratos.
Inclua o uso de suplementos anti-catabólicos em sua dieta.
Controle o hormônio do cortisol que é inimigo do anabolismo, ou seja, se o seu hormônio cortisol está alto, provavelmente você está em estado catabólico. (BlogEducaçãofísica, Tuasaude)
Assim, para evitar o catabolismo muscular é importante evitar jejum prolongado e ter uma alimentação correta e balanceada, de modo que as reações anabólicas e catabólicas fiquem equilibradas, evitando, desta forma a perda de massa muscular. Por outro lado, para evitar o catabolismo e ganhar massa magra, é importante favorecer o anabolismo.
Para favorescer o anabolismo, antes e após o treino é essencial ter uma alimentação rica em carboidratos e proteínas, pois esses compostos, por meio de reações catabólicas, são transformados em glicose (energia) e aminoácidos que são utilizados nas reações anabólicas para promover o crescimento muscular.
O crescimento muscular nada mais é do que o reparo de micro lesões que ocorrem nas fibras musculares.
O uso de suplementos que favoreçam o ganho de massa muscular, fornecem compostos mais simples e que, por meio das reações anabólicas, promovem a formação de compostos mais complexos, como as proteínas, por exemplo.
Se seu objetivo é ganho de massa magra e volume muscular, recomenda-se suplementar sua alimentação com BCAA (Branched Chain aminoacids), maltodextrina, dextrose, whey protein ou glutamina, de acordo com seu objetivo e a intensidade das atividades físicas praticadas ao longo doa semana. Esses suplementos também podem engordar se ingeridos sem fazer os exercícios.
Evite situações de estresse, pois nessas situações normalmente há aumento da concentração do hormônio cortisol no sangue, o que pode favorecer o catabolismo. Assim, é interessante realizar atividades que promovam o relaxamento, como assistir um filme, passear ao ar livre, andar de bike etc. Além disso, é importante respeitar os períodos de descanso após os treinos e dormir bem, pois assim não haverá sobrecarga muscular e haverá tempo para o organismo repor as energias necessárias para o bom funcionamento do corpo e para ser exigido novamente para o aumento do volume muscular. (Modif. de Tuasaudae)
“Como sabemos o qual é o tipo primitivo de gastrulação? O estado atual da embriologia certamente não nos permite dar uma resposta positiva a essa pergunta. Se a forma primária é a gástrula epibólica ou embólica, a plácula, a plânula delaminada unipolar ou multipolar, ou ainda um tipo diferente, está para ser visto; e o próprio fato de que a diferenciação das camadas é efetuada em tal diversidade de maneiras prova conclusivamente que esses estágios iniciais de desenvolvimento são tão suscetíveis à modificação secundária quanto os posteriores. ” (E.B. Wilson, 1892)
“How do we know what is the primitive type of gastrulation? The present state of embryology certainly does not enable us to give any positive answer to this question. Whether the primary form is the epibolic or the embolic gastrula, the plakula, the unipolar or multipolar delaminate planula, or a still different type, remains to be seen; and the very fact that the differentiation of the layers is effected in such a diversity of ways proves conclusively that these early stages of development are as susceptible to secondary modification as the later.”
(E.B. Wilson, 1892. Zoólogo e geneticista Estadunidense)
EMBRIOLOGIA
Como sabemos os organismos estão divididos em dois grupos principais, dependendo apenas se as células possuem um envoltório nuclear ou não.
Os procariotos (do grego πρό = pro: antes e κάρυον = karyon: noz, amêndoa, ou núcleo, onde estão incluídas as eubactérias e as arqueobactérias, não possuem um núcleo verdadeiro.
Os eucariotos, do grego ευ = eu : verdadeiro, bom, perfeito; e κάρυον = karyon: noz, amêndoa, núcleo que incluem os protoctistas, metazoários (animais), embriófitas (plantas) e fungos, possuem um tegumento nuclear bem formado, a carioteca, circundando seu material genético, i.e., os seus cromossomos.
Essa diferença fundamental entre os procariotos e eucariotos influencia a maneira como esses grupos organizam e utilizam seu material genético. Em ambos os grupos, a informação herdada necessária para o seu desenvolvimento e metabolismo se encontra codificada nas sequências de ácido desoxirribonucléico (DNA) dos cromossomos.
Os cromossomos procarióticos normalmente são hélices duplas de DNA, pequenas e circulares consistindo de aproximadamente 1 milhão de pares de bases.
As células eucarióticas geralmente possuem diversos cromossomos, e um simples protoctista (eucariótico) possui 10 vezes, ou mais, a quantidade de DNA encontrada na maioria dos procariotos complexos.
Além disso, a estrutura de um gene eucariótico é mais complexa do que a de um gene procariótico. A seqüência de aminoácidos de uma proteína procariótica é a reflexão direta da seqüência de DNA do cromossomo.
O DNA de um gene eucariótico que codifica uma proteína, geralmente, é dividido de tal forma que a seqüência completa de aminoácidos da proteína é derivada de segmentos descontínuos de DNA. O DNA entre os segmentos freqüentemente contém seqüências que estão envolvidas na regulação do momento e lugar em que o gene é ativado.
Cromossomos eucarióticos também são muito diferentes dos cromossomos procarióticos. O DNA eucariótico reveste complexos protéicos específicos, chamados nucleossomos, compostos por proteínas histonas.
Os nucleossomos organizam o DNA em estruturas compactas e são importantes na designação de qual gene irá se expressar em qual célula.
Nas bactérias não existem histonas. Mais ainda, células eucarióticas sofrem mitose, na qual o tegumento ou envoltório nuclear, carioteca, se parte e os cromossomos replicados são igualmente divididos entre as células filhas.
Nos procariotos, a divisão celular não é mitótica; não se desenvolve o fuso mitótico e, também, não existe tegumento nuclear para se partir. Ao invés disso, os cromossomos filhos permanecem ligados a pontos adjacentes na membrana celular, chamados mesossomos. Esses pontos de ligação são separados entre si pelo crescimento da membrana celular, e finalmente colocam os cromossomos em diferentes células filhas.
Os últimos anos do século XX encontram a biologia do desenvolvimento retornando à posição que ela ocupou no início do século: a disciplina que unifica os estudos da hereditariedade, evolução e fisiologia.
Em 1896, a primeira edição de B. Wilson do The Cell in Development and Inheritance anunciou “a verdade maravilhosa que uma única célula pode conter em seu interior sua extensão microscópica da soma-total da herança das espécies.”
Hoje, a biologia do desenvolvimento está na vanguarda desse estudo de nossa herança natural. Nos seus aspectos moleculares, ela toca a química, e a física na sua investigação dos mecanismos bioquímicos pelos quais proteínas diferentes são produzidas em células diferentes do mesmo genoma. Ela também está na liderança dos estudos evolucionários que procuram entender como mudanças macroevolucionárias ocorreram.
Ela abriu recentemente uma área nova da biologia do desenvolvimento ecológico, onde mudanças ambientais são vistas criando alterações no desenvolvimento do organismo. Durante os últimos 3 anos, a biologia do desenvolvimento também expandiu para a medicina, fundindo-se com a genética clínica para criar uma ciência revitalizada da embriologia humana, uma ciência que já se tornou importante na explanação das malformações congênitas.
EMBRIOLOGIA E EVOLUÇÃO
Segundo Gilbert (2016) as principais características do desenvolvimento animal estão ilustrados na Figura 1 abaixo.
A vida de um novo indivíduo é iniciada pela fusão do material genético de dois
gametas, o espermatozóide e o óvulo.
Essa fusão, chamada fertilização, estimula o
ovo a iniciar o desenvolvimento.
A fertilização ocorre quando o espermatozoide penetra no ovócito, e vários eventos iniciam nesse momento. Um evento importante é a fusão de dois pronúcleos, um masculino e outro feminino, levando à restauração do número diploide de cromossomos, da espécie.
No caso dos humanos, restaura-se o número de 46 cromossomos (23 cromossomos vindos do gameta masculino + os 23 cromossomos vindos do gameta feminino) o que gera a variabilidade encontrada nas espécies.
Ao final desses eventos, forma-se a célula ovo ou zigoto, e os 46 cromossomos que o formam se organizam em um fuso de clivagem.
Fertilização
Fertilização
Representação da interação entre os gametas e da reação acrossômica.
A-B) O espermatozoide liga-se à zona pelúcida e sofre a reação acrossômica;
C-D) as enzimas do acrossoma digerem a zona pelúcida, o espermatozoide faz um contato tangencial com o oócito, ocorre a fusão das membranas dos gametas, e E) o espermatozoide entra no oócito.
Os núcleos dos gametas são denominados de pronúcleos.
Para alcançar o ovócito, inicialmente, o espermatozoide passa pela corona radiata utilizando enzimas
presentes no acrossoma, como a hialuronidase, que promove a quebra de ligação das células foliculares,
dispersando-as.
A seguir, o gameta masculino se liga à zona pelúcida a qual passa a ser digerida naquele ponto pela enzima
proteolítica acrosina.
A porção anterior da membrana plasmática do espermatozóide é desprendida, assim como
as mitocôndrias são perdidas. Com a fusão das membranas plasmáticas dos gametas (ovócito e espermatozoide), o conteúdo dos
grânulos corticais do ovócito é liberado por exocitose, alterando proteínas específicas da zona pelúcida (reação
de zona) um mecanismo para evitar a poliespermia, ou seja, que outros espermatozoides fertilizem o mesmo ovócito (Nunes et alii, 2020).
A fertilização estimula o ovócito II a finalizar a segunda divisão meiótica, quando libera o segundo
corpúsculo polar e dá origem ao.
Essa célula passa a ser chamada de óvulo.
Concomitantemente, o núcleo do espermatozoide aumenta de tamanho, se descondensa, tornando-se o
pronúcleo masculino, e sua membrana se desintegra. A seguir, o centríolo do espermatozóide se coloca entre os
pronúcleos haplóides, promovendo a fusão destes ocorrendo a formação de uma nova célula diplóide, a célula ovo ou zigoto.
Na foromação dessa nova célula os envoltórios nucleares desintegram-se, e os cromossomos pareian-se. Logo apoós inicia-se a clivagem ou segmentação.
O espermatozóide pode contribuir com os centríolo(s) para organizar o fuso mitótico do zigoto. Isso ocorre, por ex., em ouriço-do-mar, onde há a entrada dos dois centríolos do espermatozoide no ovo.
Em humanos, o centríolo proximal do espermatozoide sofre duplicação no gameta feminino para formar os ásteres. Em insetos e em alguns mamíferos, como os roedores, onde há a perda da integridade dos centríolos do espermatozoide, o fuso surge a partir do centrossoma materno. (Montanari, 2013).
O gameta feminino geralmente apresenta camadas acelulares envoltórias que servem para a proteção e barreira pela qual os espermatozoides devem atravessar. Conforme a sua origem, esses envelopes pode ser classificados em primário, secundário e terciário.
Envelope primário
É secretado pelo próprio oócito. Ex.: membrana vitelínica dos cntenóforos, envelope vitelínico e camada gelatinosa em ovos de equinodermos, córion em ovos de peixe, envelope vitelínico em ovos de anfíbios, membrana vitelínica dos ovos de aves e zona pelúcida em mamíferos.
Envelope secundário
Sintetizado pelas células foliculares. Ex.: envelope vitelínico e córion em ovos de insetos.
Envelope terciário
O envelope terciário é produzido pelas células do trato reprodutor feminino, sendo adicionado ao gameta após a sua ovulação. Ex.: camada gelatinosa nos ovos de ctenóforos e de anfíbios, albúmen (clara) membranas interna e externa da casca nos répteis e aves.
Fertilização, clivagens, mórula, compactação e gástrula
Os estágios subsequentes do desenvolvimento são
coletivamente chamados de embriogênese.
A embriogênese é o processo através do qual o embrião é formado e se desenvolve. Começa no momento da fertilização, a fusão do núcleo do gameta masculino com o núcleo do gameta feminino constituirá uma nova entidade que a partir desse momento passará a chamar-se de ovo ou zigoto.
Clivagem
A segmentação ou clivagem inicia-se logo depois da fertilização pode ocorrer de muitas maneiras distintas, dependendo do tipo de ovo que cada grupo taxonômico apresenta, e está relacionada com a quantidade de vitelo presente no citoplasma o óvulo, ou seja, a quantidade de vitelo influenciará na velocidade da clivagem nos diferentes grupos animais, sendo a rapidez da clivagem, inversamente proporcional a quantidade de vitelo.
O pólo do ovo com maior quantidade de vitelo é denominado polo vegetal ou polo vegetativo. O polo oposto, é chamado de polo animal, com baixa concentração de vitelo, e é onde se encontra o núcleo da célula.
Classificação do ovo quanto à quantidade de vitelo
Desta forma podemos classificar o ovo observando a quantidade de vitelo e a forma com a qual este vitelo está distribuído, por isso, os ovos podem ser classificados em quatro grupos grandes grupos.
Ovo oligolécito ou isolécito
O ovo oligolécito ou isolécito possui pouco vitelo e este se encontra distribuído de forma relativamente homogênea no citoplasma da célula ovo.
Ovo mesolécito ou heterolécito
Ovos mesolécitos ou heterolécitos apresentam uma quantidade considerável de vitelo, que se encontra distribuído de maneira heterogênea no citoplasma. Geralmente há um lado com maior quantidade vitelo, e este lado que possui maior quantidade de vitelo é chamado de polo vegetativo, enquanto o lado com menor quantidade de vitelo é chamado de polo animal.
Ovo megalécito ou telolécito
O ovo megalécito ou telolécito apresenta uma grande quantidade de vitelo, que empurra o núcleo do gameta feminino para a região mais periférica e próxima à membrana plasmática região que é chamada de disco germinativo.
Ovo centrolécito
O ovo centrolécito apresenta quantidade considerável de vitelo, que se localiza sobretudo na região central.
Analisando-se os diferentes tipos de ovos, é possível observar dois tipos de segmentação: a segmentação holoblástica e a segmentação meroblástica.
A segmentação holoblástica pode ser ainda classificada em igual e desigual, e a segmentação meroblástica pode ser dividida em discoidal e superficial.
A segmentação holoblástica, também chamada de total, apresenta divisões em todo o ovo. Na segmentação holoblástica igual, que ocorre em ovos oligolécitos, observa-se uma divisão que forma blastômeros que apresentam relativamente o mesmo tamanho.
Na segmentação holoblástica desigual, que ocorre nos ovos mesolécitos, observa-se a formação de células pequenas (micrômeros) no polo animal e células maiores (macrômeros) no polo vegetativo (polo com maior quantidade de vitelo).
A segmentação meroblástica, também chamada de parcial, apresenta divisões apenas na região sem vitelo. Na segmentação meroblástica discoidal, que ocorre em ovos megalécitos, a divisão ocorre na região do disco germinativo.
Na segmentação meroblástica superficial, que ocorre nos ovos centrolécitos, percebe-se que a divisão (clivagem ou segmentação) acontece na região periférica do ovo.
Padrões de clivagem
O padrão de clivagem embrionária é determinado tanto pela posição dos fusos mitóticos quanto pela quantidade e distribuição de vitelo no ovo.
Uma quantidade grande de vitelo tende a inibir o clivagem, diminuindo a velicidade da mesma.
O vitelo é uma adaptação para animais que apresentam fecundação externa deenvolvimento indireto. Onde o embrião passa pela embriogênese sem receber nutrientes de fontes externas. Alguns animais, como o ouriço-do-mar, têm relativamente pouco vitelo porque se desenvolvem rapidamente em uma forma larval de natação livre que consegue nutrientes de seu ambiente.
Outros animais que apesar de apresentar fecundação interna, como marsupiais, nascem prematuramente, todavia recebem nutrição no marsúpio da mãe.
Os mamíferos placentários desenvolvem um órgão especializado (placenta) através do qual o embrião é nutrido, pela mãe, durante todo o desenvolvimento e, portanto, também possuem ovulos com pouco vitelo. Nunes et alii (2020).
Os tipos de ovos com base nas características do vitelo são descritos como:
Isolécitos, Oligolécitos ou Alécitos
Nos ovoúlos isoleticos ou oligolécitos ou alécitos, o vitelo é pouco esta esparsa uniformemente distribuído no óvulo. Por exemplo, ouriço do mar, camundongo (Equinodermos e mamífero).
Heterolécitos ou Mesolecitos
Nestes há uma quantidade moderada de vitelo, frequentemente distribuída de maneira desigual no óvulo. Ex., Anfíbios e Peixes.
Telolécitos
Neste tipo de óvulo ocorre uma grande quantidade de vitelo, concentrado no polo vegetativodo. por exemplo: Aves e repteis.
Centrolecitos
Nos ovos centrolécitos o bitelo encontra-se concentrado no meio do ovo, por exemplo ovos de insetos.
Muitos ovos são polarizados com um dos polos rico em vitelo, denominado pólo vegetativo e um polo pobre em vitelo denominado polo animal, por exemplo, sapo.
O núcleo zigótico é geralmente deslocado em direção ao pólo animal.
Zigotos com relativamente pouco vitelo (isolecitos e mesolecitos) apresentam clivagem do tipo holoblástica. O sulco de clivagem se estende por todo o ovo.
Enquanto os zigotos do tipo telolécito e centrolécitos sofrem clivagem meroblástica, onde o plano de clivagem se estende apenas à gema acumulada. Em ovos centrolécitos (muitos ovos de insetos) a clivagem é meroblástica e superficial, enquanto em ovos telolecitos (pássaros e peixes) a clivagem é discoidal.
Existem vários tipos de simetria de clivagem vistos na natureza: radial (equinodermos, anfíbios), espiral (moluscos, anelídeos), Bilateral (ascídias, tunicados), Rotacional (mamíferos). As duas figuras abaixo mostram exemplos de simetrias de clivagem holoblástica e meroblástica.
A clivagem em anfíbios é holoblástica com simetria radial, entretanto o grande volume de vitelo (óvulo mesolécito ou heterolécito) interfere na clivagem. No pólo animal, a primeira clivagem ocorre em cerca de 1 mm / min, enquanto no pólo vegetal ocorre 50-100 vezes mais devagar (0,02 mm / min). Enquanto a primeira clivagem ainda está incompleta na região vegetativa do óvulo, a segunda clivagem meridional começa a ocorrer
Clivagem num ovo de anfíbio
A terceira clivagem é equatorial, mas como os núcleos e asters estão deslocados “para os animais”, o plano de clivagem, embora perpendicular ao eixo animal vegetal, também é deslocado em direção ao pólo animal e não divide igualmente os blastômeros. O resultado são quatro blastômeros animais menores (denominados micrômeros) e quatro grandes blastômeros polares vegetais (denominados macrômeros).
Essa clivagem holoblástica desigual dá origem a um pólo animal de divisão mais rápida, composto de micrômeros menores, e a um pólo vegetal de divisão mais lenta, composto de macrômeros. O pólo animal logo é composto de muitos micrômeros pequenos e o pólo vegetal alguns macrômeros grandes cheios de vitelo. Embora a formação da blastocele comece com a primeira clivagem, ela não se torna óbvia até o estágio de 128 células.
O zigoto sofre rápidas divisões mitóticas também chamado de segmentação ou clivagem, sem que haja um aumento do volume celular, i.e., sem que haja a fase de crescimento ciclo celular.
Sucessivas divisões mitóticas do zigoto, inicia-se logo após a fecundação. e caracteriza-se por um rápido aumento do número de células por divisão mitótica. As células embrionárias resultantes são chamadas de blastômeros.
Na espécie humana este processo ocorre quando o zigoto ainda está dentro da zona pelúcida e sua divisão em blastômeros acontece cerca de 30h após sua formação.
Após o estágio de 8 células, os blastômeros mudam sua forma e se agrupam.
Esse processo é chamado de Compactação. Esse processo é provavelmente mediado por glicoproteínas de adesão.
Após o estágio de oito células, os blastômeros iniciam a compactação, por meio de moléculas de adesão e formam uma “bola” de células.
Quando possuir entre 12 a 32 blastômeros, o zigoto é chamado de mórula, que chegará ao útero próximo ao quarto dia de desenvolvimento.
Compactação
Permite maior interação célula-célula
Contribui para coesão, comunicação e a cooperação entre as células.
Pré-requisito para a segregação de células internas.
É formada por junções gap, junção de adesão e junções de oclusão que ocorrem nos blastômeros externos.
Para isto entram em cena as junções que do ponto de vita funcional podem ser classificadas em três tipos: junção de adesão, junções impermeáveis, junções de comunicação.
Junções de adesão
São também chamadas de junções aderentes, zônulas de adesão. Os hemidesmossomos e os desmossomos também são estruturas de adesão.
Junções impermeáveis
As junções impermeáveis são chamadas também de zônulas de oclusão.
Junções de comunicação
As junções de comunicação, chamadas também de junções apertadas (tight junction), junções ocludentes, junções comunicantes ou junções gap. Todas estas junções entram em atividade na construção da mórula.
Tipos de junções entre membrans plasmáticas de células adjacentes
O desenvolvimento inicia-se com a divisão do zigoto e a formação, através de mitoses sucessivas, de células chamadas de blastômeros.
Essa divisão inicial através da mitose produz células totipotentes. Essa divisão inicial é chamada e conhecida por segmentação ou clivagem.
Em humanos o estágio de de 12-32 células. Forma-se cerca de 3 dias após fecundação. Alcançando o útero cerca de 4 dias após a fertilização.
No caso dos ouriços do mar a formação da mórula é mais rápido.
Durante as rodadas de clivagem consecutivas, respectivamente de 2, 4, 8, 16, 32, 64, 128 e finalmente termina na fase de mórula. Depois de um tempo, fica difícil distinguir cada célula. O estágio pluricelular (aqui mostrado são provavelmente 128 blastômeros, figura abaixo) constituindo uma mórula. Neste estágio, o citoplasma do ovo fertilizado é completamente divido entre os vários blastômeros. Observe que esse grupo de células embrionárias tem aproximadamente o mesmo tamanho que o óvulo fertilizado original! O que ocorreu foram divisóes celulares sem o estágio de crescimento do ciclo celular.
Estágio de mórula em ouriço do mar, (A) com 32 células e (B) com 128 céulas.
Clivagem precoce em Lytechinus variegatus.ouriço-do-mar. (A) Estágio de duas células. (B1) Vista Polar do estágio inicial de quatro células. (B2) Visão lateral do estágio final de quatro células. (C) Vista Lateral do estágio inicial de oito células. (D) Visão lateral do estágio final de 16 células. Abreviaturas: as = aster; bpm = membrana plasmática do blastômero; fe = envelope de fertilização (zona pelúcida); mac = macrômeros; mes = mesômeros; mic = micrômeros; n = núcleo; sp = fuso.
Essas divisões mitóticas sucessivas e rápidas, se iniciam, no caso dos humanos, algumas horas (até 30 horas após a fecundação) e provocam um aumento do número de blastômeros (células) rapidamente.
A rapidez da clivagem é inversamente proporcional à quantidade de vitelo presente no ovo. Quanto maior maior a quantidade de vitelo mais lento ocorrerá a clivagem e quanto menor a quantidade de vitelo maior será a velocidade da clivagem e todo o processo.
As células resultantes da clivagem são chamadas de blastômeros. A cada divisão formam-se blastômeros menores.
Quando existirem oito blastômeros, os mesmos mudam sua forma e se unem, no processo de compactação, formando uma estrutura compacta de células. Essa proximidade das células permite a formação da massa celular interna (MCI).
Formação do blastocisto de camundongo. No estágio de oito células, o embrião se compacta para produzir uma mórula (A). Após a próxima divisão celular, o embrião começa a produzir uma cavidade ou blastocele e forma um blastocisto no estágio de 32 células (B). Duas populações de células emergem durante a formação do blastocisto. As células internas da mórula (brancas) tornam-se a massa celular interna (ICM), um grupo de células-tronco pluripotentes excentricamente posicionadas dentro do blastocisto. As células da camada externa da mórula (cinza) formam o primeiro epitélio, o trofectoderma, que se diferencia nas linhagens trofoblásticas da placenta. O blastocisto se expande por acúmulo de líquido adicional dentro da blastocele e eclode livre da zona pelúcida (C). No camundongo, as células trofectoderme polares em contato com o MCI proliferam, enquanto as células trofectoderma murais restantes se diferenciam em células trofoblásticas gigantes. (Armant, D.R.2006).
Formation of the mouse blastocyst. At the eight-cell stage, the embryo compacts to produce a morula (A). After the next cell division, the embryo begins to produce a cavity or blastocoel and forms a blastocyst by the 32-cell stage (B). Two populations of cells emerge during blastocyst formation. The inside cells of the morula (white) become the inner cell mass (ICM), a group of pluripotent stem cells eccentrically positioned within the blastocyst. Cells in the outer layer of the morula (gray) form the first epithelium, the trophectoderm, which differentiates into the trophoblast lineages of the placenta. The blastocyst expands by accumulating additional fluid within the blastocoel and hatches free of the zona pellucida (C) . In the mouse, polar trophectoderm cells contacting the ICM proliferate, while the remaining mural trophectoderm cells differentiate into trophoblast giant cells.
Desenvolvimento embrionário normal de embriões de bovinos.
1) Não fertilizado, 2) duas (2) células a 12 células, 3) Mórula precoce, 4) Mórula,
(a) Comparison of developmental stages of in vivo-derived bovine embryos collected on days 6–8 after estrus. At day 7, a stage code 1 embryo is considered an unfertilized ovum and a stage code 2 embryo is considered dead or degenerate. (b) Expanded hatched blastocyst typically recovered on or after day 8 after estrus; note the empty zona pellucida on the right and the expanded embryo on the left. (Jahnke, West & Young, 2010)
Por volta de três dias após a fecundação forma-se a mórula, que é o primeiro estágio do desenvolvimento.
A mórula é constituída de 12 a 32 blastômeros, sendo formada por uma camada interna que é chamada de massa celular interna, e uma camada externa é circundada por uma outra camada, que é a camada celular externa.
Por todo reino animal (reino metazoa) existe uma incrível variedade de tipos embrionários, mas a maioria dos padrões de embriogênese compreende variações em quatro temas:
1 Tema.
Ocorrência de clivagem imediatamente após a fertilização e formação da mórula.
Clivagem é uma
série de divisões mitóticas extremamente rápidas, onde o enorme volume citoplasmático do zigoto é dividido em numerosas células menores. As céluls se dividem sem que haja a fase de crescimento do ciclo dcelular. Desta fomr aos blastômeros resultantes são cada vez menores.
Clivagem em ouriço do mar
Clivagem e formação da mórula
Essas células
são chamadas blastômeros e, ao fim da clivagem, eles geralmente formam uma
esfera maciça conhecida como mórula.
A compactação celular que se observa facilita a comunicação entre as células da mórula. Então, podemos dizer que aclivagem cria a multicelularidade.
A informação espacial desempenha um papel crítico na formação do blastocisto. Durante o desenvolvimento pré-implantação, o trofectoderma se diferencia das células que revestem a parte externa da mórula compactada (Fig. 1). Pistas de desenvolvimento obtidas pela presença ou ausência de contatos celulares assimétricos determinam o destino da célula após a compactação (1). As células externas se diferenciam em trofectoderma, o primeiro epitélio e precursor de todas as linhagens trofoblásticas, enquanto as células internas tornam-se a massa celular interna pluripotente (ICM). Durante a diferenciação do trofectoderma, junções estreitas se formam entre os blastômeros externos e eles começam a transportar ativamente o fluido para o embrião, gerando uma blastocele.
À medida que o blastocisto se forma, o trofectoderma é alongado e o agrupamento de células que forma o MCI fica confinado a um setor da parede do trofectoderma (2). O trofoblasto se origina da camada trofectoderme do blastocisto durante o desenvolvimento da peri-implantação. Portanto, muito pode ser aprendido sobre o desenvolvimento de trofoblasto por meio da cultura de blastocistos in vitro. O blastocele separa fisicamente algumas das células do trofectoderma do ICM e, assim, fornece pistas espaciais para a diferenciação do trofectoderma mural. A trofectoderme polar compreende as células externas que contatam diretamente a ICM (3). Sob a direção da ICM, as células da trofectoderme polar proliferam rapidamente até se deslocarem para dentro da trofectoderme mural, onde a divisão celular diminui consideravelmente (4,5).
No camundongo, as células trofectoderme mural endoreduplicam seu DNA e se diferenciam em células trofoblásticas gigantes invasivas.
Estrutura do blastocisto
1. A camada que o reveste, chamada de trofectoderma. Suas células vão originar o trofoblasto, responsável pela implantação do embrião no útero, por sua nutrição e pelo desenvolvimento da placenta e os outros anexos fetais.
2. A massa celular interna (MCI), o embrioblasto, com suas células agregadas em um polo do embrião. Esse conjunto de células vão dar origem ao embrião em si.
3. A blastocele, uma cavidade contendo líquidos nutritivos para o embrião e que faz parte da formação dos celomas embrionários (Soares, 2020).
Morphogenetic behavior of the micromere-PMC lineage during early sea urchin embryo development (Schoenwolf, 2001). (A) Micromeres (red) are formed in the fourth, unequal cleavage. (B) Subsequent divisions result in 32 PMCs at the vegetal pole of the hollow blastula. (C) PMCs then ingress into the mesenchyme blastula (MB) in an epithelialmesenchymal transition. (D and E) PMCs show directional migration, forming two ventrolateral clusters (VLCs) and a stereotypical pattern of strands in the late gastrula (LG). Cell-cell fusion leads to the formation of two syncytial masses. Synthesis of the endoskeleton inside each syncytium commences with the formation of a calcareous granule that grows into the triradiate spicule rudiments. (Moreno et alii, 2019).
Schematic overview of sea urchin development showing the origin and
fate of selected cell populations. (A) Fertilized egg or zygote, hemisection. (B) 16-
cell stage, surface view. (C) 64-cell stage, surface view. (D) Mesenchyme blastula or
early gastrula, hemisection. (E) Late Gastrula, hemisection. (F) Prism, hemisection.
(G) Early pluteus larva, hemisection. Light dots = ectoderm. Heavy
dots = endoderm, coelomic mesoderm, and secondary mesenchyme.
Stippling = primary mesenchyme.
Abbreviations: a = animal pole; an = anus; an1 = first tier of animalized
blastomeres; an2 = second tier of animalized blastomeres; ar = archenteron;
at = apical tuft of cilia; bc = blastocoel; bp = blastopore; br = body rod; ci= cilia;
es = esophagus; in = intestine; m1 = primary mesenchyme; m2 = secondary
mesenchyme; mac = macromeres; mes = mesomeres; mic = micromeres;
mo = mouth; poa = postoral arm; por = postoral rod; pra = preoral aram;
prr = preoral rod; sp = spicule; sr = skeletal rod; st = stomodaeum; sh = stomach;
tr = transverse rod; v = vegetal pole; vegl = first tier of vegetalized blastomeres;
veg2 = second tier of vegetalized blastomeres.
prr = preoral rod; sp = spicule; sr = skeletal rod; st = stomodaeum; sh = stomach;
tr = transverse rod; v = vegetal pole; vegl = first tier of vegetalized blastomeres;
veg2 = second tier of vegetalized blastomeres. (Conway. C; Conway. A, Igelsrud, s/d).
QUAL A FUNÇÃO DO BLASTOCELE?
A blastocele separa espacialmente as células para que não se toquem. As células no teto do blastocele normalmente se tornam ectoderma. Se você transplantar células do teto da blastocele próximo às células vitelinosas na base da blastocele, elas se diferenciarão como mesoderme. Os derivados mesodérmicos são normalmente produzidos a partir de células adjacentes aos precursores endodérmicos.
Uma possibilidade que exploramos a fundo é que as células do polo vegetativo “induzam”, por meio de interações célula-célula, as células adjacentes a se tornarem mesodérmicas. Assim, a formação do blastocele pode ser necessária para evitar interações "indutivas" inapropriadas entre as células iniciais do blastocisto.
A segunda necessidade óbvia para a blastocele pode ser durante o estágio subsequente de desenvolvimento, gastrulação, onde as células migram para o interior da blastocele e se diferenciam. (Bastiani)
Os mamíferos, por possuírem placenta que lhes fornece alimento e oxigênio durante a gestação, possuem ovo do tipo oligolécito, em que há vitelo escasso, sendo este distribuído de forma homogênea. Desta forma, a clivagem é do tipo holoblástica, i.e., o sulco de clivagem ocorre por todo o ovo.
O ovo dos mamíferos sofre clivagens lentas durando, cada divisão, cerca de 12 a 24 horas; a primeira ocorre enquanto o zigoto caminha pela tuba uterina em direção ao útero.
Quanto ao plano de divisão, essa primeira divisão é meridional e origina dois blastômeros.
A segunda divisão, uma célula (blastômero) se divide meridionalmente enquanto a outra, equatorialmente, para dar origem a quatro blastômeros, ou seja, tem padrão rotacional. Portanto, mamíferos possuem ovos oligolécitos com clivagem do tipo holoblástica rotacional. (Nunes et alii, 2020)
2 Tema.
Seguem as contínuas divisões e reorganização celular, ficando uma pequena população dos blastômeros mais internamente localizada, chamada de MCI ou massa celular interna, circundada por uma camada externa de várias células que constituirão a trofecderme ou blastoderme (torfectoderme). Por um processo denominado cavitação, há o surgimento de uma cavidade interna cheia de líquido, denominada de blastocele. Esse estágio é chamado de blástula, ou blastocisto.
Após a redução na taxa de divisão mitótica, os blastômeros passam por mudanças dramáticas quanto às suas posições, um em relação ao outro. Essa série de redistribuição de células é chamada de gastrulação. Como resultado da gastrulação, o embrião típico contém três regiões celulares chamadas camadas germinativas ou folhetos germinativos*. O ectoderma, a camada exterior, o endoderma, camada interior, e o mesoderma, a camada intermediária.
*Do Latim germen, significa “broto” ou “rebento” (a mesma raiz da palavra germinação). Os nomes dos três folhetos germinativos são do Grego:ectodermade ektos (fora) mais derma (pele); mesoderma de mesos (meio) e endoderma de endon (dentro).
O ectoderma, a camada exterior, produz as células da epiderme e do sistema nervoso.
O endoderma, camada interior, produz o revestimento do sistema digestório (tubo digestivo) e órgãos associados (pâncreas, fígado, pulmões, etc.).
E o mesoderma, camada intermediária (do meio), dará origem a diversos órgãos
3) Uma vez que as três camadas embrionárias estão estabelecidas, as células
interagem umas com as outras e se reorganizam para produzir tecidos e órgãos.
Esse processo é chamado organogênese. Nos cordados em geral e nos vertebrados, a organogênese é iniciada quando uma série de interações celulares induzem as células ectodérmicas da porção mediana do dorso a formar o tubo neural oco dorsal. Esse tubo originará o cérebro (sistema nervoso) e a medula espinhal. Muitos órgãos contêm células de mais de um folheto ou camada embrionária, e não é incomum o exterior de um órgão ser derivado de um determinado folheto e o interior de outro. Também durante a organogênese, algumas células sofrem longas migrações de seu lugar de origem até sua localização final.
Essas células migrantes incluem os precursores das células sangüíneas, células linfáticas, células pigmentadas e gametas.
A maior parte dos ossos de nossa face são provenientes de células que migraram ventralmente da região dorsal da nossa cabeça.
4. Como observado na Figura 1.1, em muitas espécies, uma parte especializada
do citoplasma do ovo dá origem às células que são precursoras dos gametas.
Essas células são chamadas de células germinativas, sendo destinadas à função reprodutiva. Todas as outras células do corpo são chamadas células somáticas. Essa separação entre células somáticas (que dão origem a um corpo individual) e células germinativas (que contribuem para a formação de uma nova geração) é freqüentemente uma das primeiras diferenciações que ocorrem durante o desenvolvimento animal. As células germinativas finalmente migram para as gônadas, onde se diferenciam em gametas. O desenvolvimento de gametas, chamado de gametogênese, normalmente não é completado até que o organismo tenha se tornado fisicamente maduro. Na maturidade, os gametas podem ser liberados e participar de uma fertilização dando início a um novo embrião. O organismo adulto finalmente sofre envelhecimento e morre.
Blástula ou blastocisto (esquerda esquema mostrando as partes, a direita foto colorida artificialmente). (Fonte: bastiani.biology)
O trofoblasto é camada celular externa do blastocisto, contribui somente para formação da placenta.
Embrioblasto
Conjunto de células internas (MCI, massa celular interna) que contribuem para formação do embrião e das membranas extraembrionárias O embrioblasto forma o disco bilaminar composto de:
Epiblasto
O epiblasto é conjunto celular mais interno do embrioblasto.
Hipoblasto
O hipoblasto constitui a camada celular inferior do embrioblasto, somente participará na formação de membranas extraembrionárias (saco vitelínico).
Na nidação ocorre a penetração gradual do blastocisto no endométrio, a proliferação do citotrofoblastoe transformação em sincíciotrofoblasto, o embrioblasto forma disco bilaminar: epiblasto e hipoblasto, e ocorre o aparecimento da cavidade amniótica por cavitação no epiblasto.
O endométrio (decídua), onde o blastocisto irá nidar, é a camada interna do útero, rica em glândulas e vasos sanguíneos, prolifera durante ciclo ovariano representa contribuição materna da placenta.
Citotrofoblasto: camada celular proliferativa do trofoblasto.
Sincíciotrofoblasto: camada externa do trofoblasto, altamente ativa em síntese protéica (enzimas proteolíticas); cresce por meio do citotrofoblasto, células perdem separação por membranas e tornam-se sinciciais, i.e. citoplasma comum multinucleado. O citotrofoblasto e sincíciotrofoblasto são os elementos embrionários da placenta.
Blastocisto implantado no endométrio (9dias)
Implantação completa do blastocisto no endométrio, formação de lacunas no sincíciotrofoblasto, a expansão do hipoblasto sobre face interna do trofoblasto resulta no saco vitelínico primário, absorção da secreção de glândulas uterinas.
Âmnio
O âmnio é uma membrana extraembrionária que delimita a cavidade ou câmara amniótica cuja função é a proteção contra desidratação e proteção contra choques mecânicos. Constitui-se em uma adaptação ao desenvolvimento embrionário em condições terrestres em vertebrados. O âmnio surgiu nos répteis cujos embriões se desenvolviam fora do corpo materno, dentro do ovo âmniótico-alantoidiano ou reptiliano. Animais que possuem âmnio são chamados de amniotas e aqueles que não possuem âmnio são chamados de anamniotas.
Cavidade amniótica
É a câmara protetora do embrião, preenchida com líquido amniótico.
Saco vitelínico
O saco vitelínico é primeiro anexo embrionário a aparecer. É gerado a partir do hipoblasto (em aves e répteis e mamíferos monotremadas (aplacentários) o saco vitelínico contem grande quantidade de vitelo. Em mamíferos placentários (Eutheria), apresenta muito pouco vitelo ou sem vitelo. Representa um vestígio evolutivo, com funções importantes na formação do embrião.
Embrião de 12 dias
Enzimas do sincíciotrofoblasto abrem vasos sanguíneos maternos, sangue flui para lacunas do trofoblasto. Ocorre a expansão de mesoderma extraembrionário, espaço entre citotrofoblasto e saco vitelínico é preenchido por mesoderma extraembrionário. Ocorre a formação de cavidades (celomas) no mesoderma extraembrionário.
Córion (placa coriônica)
Revestimento externo dos celomas extraembrionários, colado internamente ao trofoblasto; passa por vascularização.
Pedúnculo placentar
É o pedúnculo de mesoderma extraembrionário condensado no polo posterior ao epiblasto, cuja função é fixar o embrião á placenta e formar o futuro cordão umbilical.
Alantóide
O alantoide é uma bolsa que surge do endoderma extraembrionário. Logo, seu exterior é revestido de mesoderma e, tal como a vesícula vitelina, está ligado ao intestino do embrião. A função do alantoide é armazenar as excretas.
Os excretas são resíduos metabólicos das substâncias produzidas durante o desenvolvimento do embrião. O alantóide invade o pedúnculo e conecta-se com placa coriônica (córion-alantóide).
Em aves desenvolve-se como broto do intestino posterior que se expande no celoma extraembrionário. É importante no processo da vascularização do trofoblasto, vasos da circulação embrionária na placenta.
Função excretora do alantóide. Em embriões de répteis e aves é no alantóide que são armazenados os produtos da excreção nitrogenada, representados sobretudo pelo ácido úrico, substância esbranquiçada e pouco solúvel em água, menos tóxica que a amônia dos peixes e a uréia dos mamíferos.
Não é o nascimento, casamento ou a morte mas a gastrulação o tempo mais importante de sua vida. (Lewis Wolpert) (Shaw s/d).
Gastrulação em organismos protostômios e deuterostômios
Gastrulação é o processo de movimentos de células e tecidos altamente coordenados pelo qual as células da blástula são rearranjados e ganham novas posições, estruturando um plano corpóreo de duas ou três camadas do organismo.
É o processo pelo qual o disco embrionário bilaminar se transforma em trilaminar. A gastrulação é o início da morfogênese.
Ocorre durante a 3ª semana do desenvolvimento humano.
Durante a gastrulação são formados a linha primitiva, as camadas germinativas ou folhetos embrionários a notocorda.
A notocorda define o eixo primitivo do embrião, forma o tubo neural dorsl oco, serve
de base para a formação dos ossos da coluna vertebral, da cabeça, o esqueleto axial, e indica o local dos futuros corpos
vertebrais.
A linha primitiva estabelece o plano básico do corpo
nos vertebrados: eixo central (linha média), simetria
bilateral, superfícies ventrais e dorsais e extremidades
cefálica e caudal. O nó primitivo ou nó de Hensen define a direção cefálica.
Este processo determina os eixos anteroposterior (AP), dorsoventral (DV), e esquerdo-direito (ED) e também especifica as camadas ou folhetos germinativos, ectoderme, mesoderme e endoderme (nos organismos triblásticos), as quais darão origem a diferentes órgãos e tecidos.
Durante a gastrulação células do ectoderma
migram pelo nó e fosseta primitiva formando o
endoderma, notocorda e o mesoderma intra-embrionário.
A ectoderme é a camada mais externa e dá origem a epiderme e seus anexos (pelos e glândulas) e ao sistema nervoso (cérebro, medula, nervos e gânglios), mucosa bucal, mucosa anal, mucosa vaginal e esmalte dos dentes.
A mesoderme dá origem ao sistema esquelético, muscular, circulatório e urinário, cartilagens, sangue, vasos sanguíneos, dentina e polta dos dentes.
A endoderme, é a camada mais interna, dá origem ao sistema digestório e anexos, aos revestimentos epiteliais das passagens respiratórias e do trato gastrointestinal, incluindo as glândulas desse trato e as células glandulares dos órgãos associados, como fígado e pâncreas.
Movimentos celulares durante a gastrulação
Os movimentos celulares que ocorrem durante a gastrulação são gerados por alterações do citoesqueleto e da afinidade de cada célula com a lâmina basal e com células adjacentes.
Eles envolvem o embrião inteiro e, as migrações celulares em uma parte do organismo gastrulante, devem estar intimamente coordenadas com outros movimentos ocorrendo simultaneamente.
Mesmo que o padrão de gastrulação seja extremamente variado em todo o reino animal, relativamente poucos mecanismos estão envolvidos. A gastrulação, geralmente, envolve os seguintes tipos de movimentos:
Embolia ou Invaginação
O dobrar para dentro de uma região de células, de maneira semelhante à cavidade formada quando se empurra com o dedo a superfície de uma bola de borracha macia.
Involução
A internação ou movimento de interiorizarão de uma camada externa em expansão, de modo a se espalhar na superfície interna das células externas remanescentes.
Ingressão
A migração de células individuais da camada superficial para o interior do embrião.
Delaminação
A separação de uma camada celular em duas ou mais camadas mais ou menos paralelas.
Epibolia
O movimento de camadas epiteliais (usualmente de células ectodérmicas) que se espalham como uma unidade e não individualmente, para envolver as camadas mais profundas do embrião.
Gastrulation, Mesoderm Formation, and Neurulation in the Lancelet Amphioxus, a Non-vertebrate Chordate. In these classic descriptions (by Edwin Grant Conklin and Berthold Hatschek), the formation of tissue layers in a non-yolky chordate egg is clearly seen. Bottom row depicts sections of embryos cut at right angles to those in the top row. (Gastrulation)
NEURULAÇÃO
ORGANOGÊNESE
Nêurula
Gastrulação
Movimentos celulares que estabelecem os folhetos embrionários no início da embriogênese:
Ectoderma
Folheto embrionário mais externo gera epiderme, tubo neural dorsal, sistema nervoso central e derivados da crista neural
Mesoderma
Folheto embrionário intermediário, gera esqueleto cartilaginoso e ósseo, musculatura, tecidos conjuntivos, sistemas cardiovascular e urogenital.
Endoderma
Folheto embrionário mais interno gera revestimento interno do trato intestinal e participa de órgãos anexos, pulmão, fígado, pâncreas.
NEURULAÇÃO
A neurulação é o processo que envole a formação da placa neural e das pregas neurais e o fechamento dessas pregas para formar o tubo neural dorsal oco que dará origem ao sistema nervoso.
Formação da placa neural e tubo neural dorsal.
A notocorda em desenvolvimento induz a formação
da placa neural.
No 18º dia inicia a invaginação da placa neural formando
um sulco neural mediano, com as pregas neurais de
ambos lados (proeminentes na região cefálica, primeiros sinais de desenvolvimento do encéfalo.
No fim da terceira semana ocorre aproximação das pregas neurais,
que começam a fundir-se, formando o tubo neural dorsal oco.
Nos humanos A formação do tubo neural começa no início da
4ª semana (dias 22 a 23) e termina no final da 4ª
semana, quando ocorre o fechamento do
neuróporo caudal (posterior).
O tubo neural se fecha primeiramente na região
medial do embrião.
As extremidades abertas são os neuróporos
rostral e caudal.
O neuróporo rostral se fecha no dia 25 e o caudal no dia 27.
O fechamento dos neuróporos coincide com o estabelecimento da
circulação vascular no tubo neural.
Formação da placa neural
Embrião humano de 17 dias
Neuroepitélio do tubo neural: Zona ventricular: origina todas as células nervosas (ficam células neuroepiteliais indiferenciadas)
Zona intermediária: entre a zona ventricular e marginal (células neuroepiteliais diferenciadas em neuroblastos)
Zona marginal: composta da parte externa das células neuroepiteliais (axônios).
Origem das células neurais
Os neurônios unipolares dos gânglios espinhais derivam das células da crista neural. Inicialmente são bipolares, depois seus prolongamentos se unem, formando um T.
O processo periférico dos gânglios espinhais vão para as terminações sensitivas das estruturas somáticas ou viscerais.
O processo central penetram na medula espinhal e constituem as raízes dorsais dos nervos espinhais.
Na neurulação ocorre a formação e fechamento das pregas neurais e formação de segmentos, chamados de somitos que ocorre em paralelo no ectoderma (tubo neural dorsal) e no mesoderma paraxial (somitos).
Fechamento das pregas neurais e formação dos somitos
As paredes do tubo neural se espessam reduzindo o espaço interno do tubo gradualmente até restar o canal central da medula espinhal.
O espessamento diferencial da medula espinhal, produz o sulco limitante. Este sulco separa a parte dorsal, a placa (lâmina) alar, da parte ventral, a placa (lâmina) basal.
Os corpos celulares das placas alares formam os cornos dorsais (cinzentos), núcleos aferentes, sensitivos.
Os corpos celulares das placas basais formam as colunas cinzentas, ventrais, e laterais, cornos ventrais e laterais. Os axônios do corno ventral formam as raízes ventrais do corno espinhal, eferentes, motoras.
Os corpos celulares das placas alares formam os cornos dorsais (cinzentos), núcleos aferentes (sensitivas). Os corpos celulares das placas basais formam as os cornos ventrais e laterais fibras eferentes (motoras).
Crista neural vs mesoderma. Origem embrionária do crânio ósseo em cinco organismos modelo de vertebrados, dispostos em uma filogenia de vertebrados simplificada.
Territórios derivados de cristas neurais (azul) foram verificados experimentalmente em cada espécie, embora as contribuições específicas de correntes migratórios individuais sejam relatadas apenas para galinha, axolote e Xenopus. A derivação de componentes remanescentes do mesoderma (cor magenta) foi verificada experimentalmente em camundongos e galinhas e é presumida para as demais espécies. As pontas de seta apontam para a interface crista neural-mesoderma no teto do crânio, que é deslocada caudalmente em Xenopus. Os dados para o peixe-zebra são das refs 8, 9; diagrama é baseado na ref. 8 (figura reproduzida com permissão da PLoS). Os dados para axolotl e Xenopus são do presente estudo; os crânios são redesenhados das referências 16, 42, respectivamente (figuras reproduzidas com permissão de John Wiley and Sons). Os dados para frango são da ref. 43; diagrama é baseado na ref. 44 (figura reproduzida com permissão de John Wiley and Sons). Os dados para o camundongo são das referências 7, 45; diagrama é baseado em refs 4, 46 (figura reproduzida com permissão de John Wiley and Sons). F, frontal; Fp, frontoparietal; N, nasal; P, parietal; Px, pré-maxila; quadrado, esquamosal. (Piekarski, Gross & Hanken, 2014; Piekarski, Gross & Hanken, 2014).
Neural crest vs mesoderm. Embryonic origin of the bony skull in five vertebrate model organisms arrayed on a simplified vertebrate phylogeny.
Neural crest-derived territories (blue) have been verified experimentally in each species, although the specific contributions from individual migratory streams are reported only for chicken, axolotl and Xenopus. Derivation of remaining components from mesoderm (magenta) has been verified experimentally in mouse and chicken and is presumed for the remaining species. Arrowheads point to the neural crest–mesoderm interface in the skull roof, which is displaced caudally in Xenopus. Data for zebrafish are from refs 8, 9; diagram is based on ref. 8 (figure reproduced with permission from PLoS). Data for axolotl and Xenopus are from the present study; skulls are redrawn from refs 16, 42, respectively (figures reproduced with permission from John Wiley and Sons). Data for chicken are from ref. 43; diagram is based on ref. 44 (figure reproduced with permission from John Wiley and Sons). Data for mouse are from refs 7, 45; diagram is based on refs 4, 46 (figure reproduced with permission from John Wiley and Sons). F, frontal; Fp, frontoparietal; N, nasal; P, parietal; Px, premaxilla; Sq, squamosal. (Piekarski, Gross & Hanken, 2014).
As regiões coloridas representam contribuições para o osteocrânio dos três fluxos migratórios CNC em quatro sistemas de modelos de tetrápodes. As contribuições em nível de fluxo (corrente) não são conhecidas no camundongo, mas presume-se que sejam semelhantes às do frango, 46. É muito parcimonioso postular que urodelos e amniotas compartilham um padrão comum de derivação CNC, que evoluiu não depois de seu ancestral tetrápode comum (barra azul na filogenia simplificada), e que o padrão único em Xenopus evoluiu após o clado de anuros divergir de urodeles (barra verde). (Piekarski, Gross & Hanken, 2014).
Coloured regions depict contributions to the osteocranium from the three CNC migratory streams in four tetrapod model systems. Stream-level contributions are not known in the mouse, but they are presumed to resemble those in the chicken 46. It is most parsimonious to posit that urodeles and amniotes share a common pattern of CNC derivation, which evolved no later than their common tetrapod ancestor (blue bar on the simplified phylogeny), and that the unique pattern in Xenopus evolved after the anuran clade diverged from urodeles (green bar).(Piekarski, Gross & Hanken, 2014).
As regiões coloridas denotam contribuições de fluxos migratórios individuais de CNC. Os rótulos vermelhos denotam ossos homólogos que têm uma origem embrionária diferente entre as espécies. Os dados para axolotl e Xenopus, dois anfíbios, são do presente estudo; os crânios são redesenhados das referências 16, 42, respectivamente. Os dados para a galinha doméstica, uma amniota, são da ref. 43; diagrama é baseado na ref. 44 (figura reproduzida com permissão de John Wiley and Sons).(Piekarski, Gross & Hanken, 2014).
À medida que o tubo neural está se fechando, certas células se dissociam dele e ficam entre o tubo neural e a epiderme sobrejacente. Estas são conhecidas como células da crista neural (Figura 38.16). São células em forma de estrela que são chamadas de “pluripotentes” porque podem se diferenciar de muitas maneiras diferentes. As células da crista neural migram extensivamente e têm incríveis capacidades de desenvolvimento. Eles não apenas formam neurônios sensoriais e partes importantes do sistema nervoso autônomo, mas também são responsáveis pela maioria dos ossos do crânio, células pigmentares e muitas outras estruturas. Sem as células da crista neural, os vertebrados não teriam cabeças, como as reconhecemos.
Coloured regions denote contributions from individual migratory streams of CNC. Red labels denote homologous bones that have a different embryonic origin between species. Data for axolotl and Xenopus, two amphibians, are from the present study; skulls are redrawn from refs 16, 42, respectively. Data for the domestic chicken, an amniote, are from ref. 43; diagram is based on ref. 44 (figure reproduced with permission from John Wiley and Sons).
As the neural tube is closing, certain cells dissociate from it and come to lie between the neural tube and the overlying epidermis. These are known as neural crest cells (FIGURE 38.16). They are star-shaped cells that are termed “pluripotent” because they can differentiate in many different ways. Neural crest cells migrate extensively and have amazing developmental capacities. Not only do they form sensory neurons and major parts of the autonomic nervous system, but they are also responsible for most of the skull bones, the pigment cells, and many other structures. Without neural crest cells, vertebrates would not have heads, as we recognize them.
Neuruação e diferenciação do mesoderma em vertebrados.
Figura 38.16: Neurulação e Diferenciação do Mesoderma em Vertebrados (A) Esta ilustração de um ovo de rã mostra como as pregas neurais se unem e se fecham, liberando células da crista neural. O mesoderma estende-se entre o ectoderma e o endoderma. As três divisões do mesoderma do tronco: somito, mesoderma intermediário e mesoderma da placa lateral, são aparentes. (B) No início da neurulação dos vertebrados, o ectoderma da placa neural é plano. A placa neural invagina e se dobra, formando um tubo. As células da crista neural são liberadas. (C,D) Micrografia eletrônica de varredura do tubo neural em um pintinho, antes e após o fechamento.
What two major mechanisms are responsible for determining the differentiated state of a cell?
What is the role of the notochord in neurulation?
A chordate synapomorphy (a shared derived trait) is a hollow dorsal nerve chord (see Concept 23.5). How does the process of neurulation explain why the spinal cord has a central cavity?
What major tissues does ectoderm form? What three major mesodermal regions form in chordates, and what does each form?
Piekarski, N. Gross, J.B. Henken, J. Evolutionary innovation and conservation in the embryonic derivation of the vertebrate skull. Nat. Commun. 5:5661 doi: 10.1038/ncomms6661 (2014).
Entre os diversos tipos de desnutrição, as deficiências protéicas estão entre as formas mais preocupantes. A qualidade protéica de um alimento está relacionada com sua composição de aminoácidos, principalmente os essenciais, e com sua digestibilidade (Ferreira et al., 2005). Em geral, os alimentos de origem animal apresentam melhor qualidade protéica quando comparados às fontes de alimentos vegetais. Algumas pesquisas revelaram que as fontes de proteínas vegetais fornecem 65% do total de proteínas ingeridas no mundo inteiro, com grãos de cereais representando 47%. Além disso, constatou-se também, que os alimentos de origem animal são a principal fonte de proteínas para humanos em países desenvolvidos, enquanto que nos países pobres ou em desenvolvimento, os alimentos de origem vegetal são as fontes mais importantes de proteínas para a população (Millward et al., 1999). No entanto, alguns grupos vegetais, como os cereais, apresentam deficiências de lisina e treonina, enquanto que as leguminosas apresentam deficiência no aminoácido metionina (Azevedo et al., 2006).
Os aminoácidos são sintetizados nos vegetais em complexas vias metabólicas controladas por enzimas, substratos e pelos próprios aminoácidos que são os produtos finais. Além dos vinte aminoácidos que são comumente produzidos e incorporados nas proteínas, mais de trezentos tipos adicionais foram identificados nos vegetais (Ferreira et al., 2005).
Os aminoácidos desempenham importantes funções como transportadores de nitrogênio para diferentes partes nos vegetais (Azevedo et al., 2006); reguladores em diversos processos envolvidos em resposta a diferentes condições ambientais e sendo, ainda, importantes para a qualidade nutricional das proteínas presentes nas sementes (Azevedo et al., 2006). Entre os vinte aminoácidos incorporados nas proteínas, nove (lisina, treonina, metionina, fenilalanina, triptofano, isoleucina, leucina, valina e histidina) são denominados essenciais porque não são sintetizados por humanos e animais monogástricos, sendo adquiridos nas dietas alimentares (Ferreira et al., 2005) (Berdejo & Azevedo s/d).

















































































































































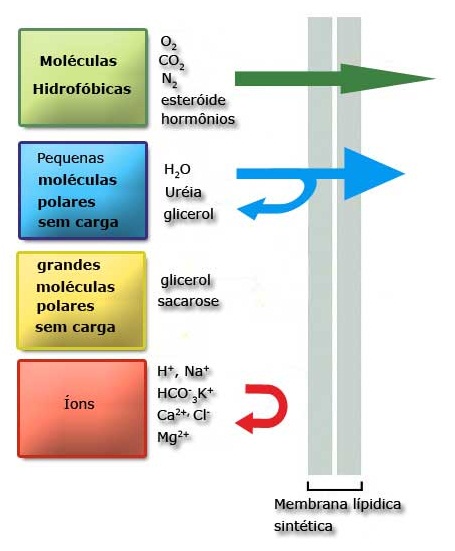
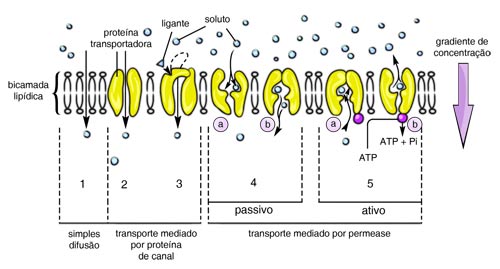

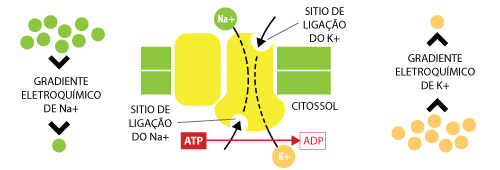




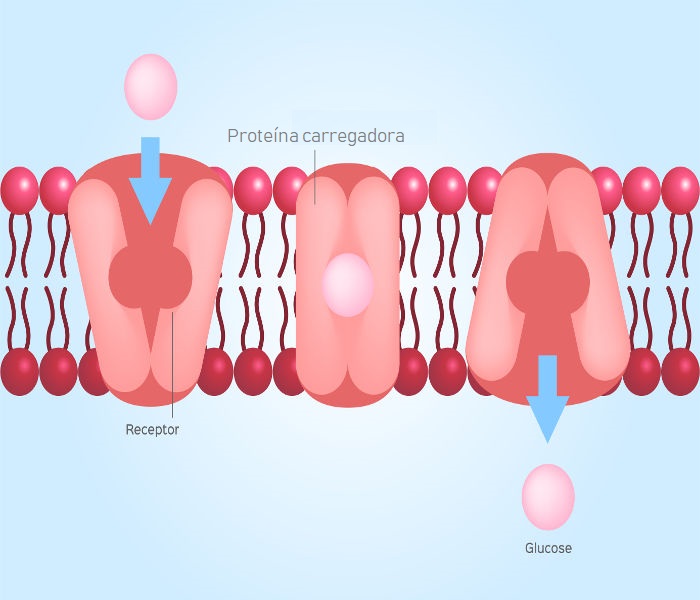

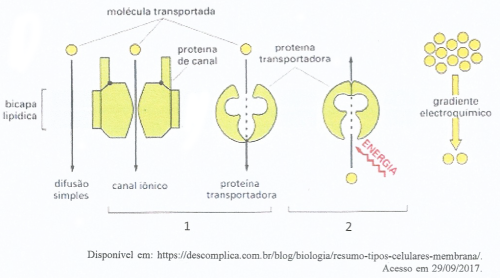
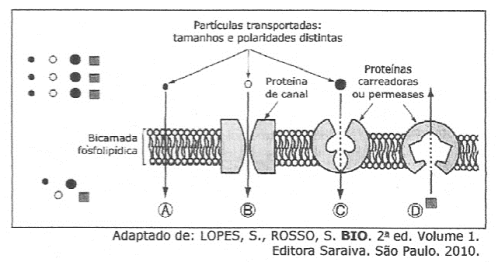































































































{kind=link}




0 comentários:
Postar um comentário