FILO CHORDATA
Filo dos CORDADOS
CARACTERÍSTICAS E SINAPOMORFIAS

Formação da notocorda
(copie esta imagem e cole no paintbrush para vê-la em tamanho grande)







Formação da notocora e dos mesodermas (paraxial, intermediário e lateral)



Neurulação eem anfíbios

Ver


FILO CHORDATA
CARACTERÍSTICAS, ORIGEM E EVOLUÇÃO
O nome do filo Chordata, vem do grego: νωτοχορδή = νωτιαία = notos, dorso, costas + χορδή = chorde: corda, cordão, (νωτοχορδή, νωτιαία χορδή).
A notocorda é uma estrutura embrionária comum a todos os organismos do filo Chordata, fornecendo pistas mecânicas e de sinalização química para o embrião em desenvolvimento. Em vertebrados, a notocorda surge do organizador dorsal e é crítica para o desenvolvimento adequado dos vertebrados. Essa estrutura evolutiva conservada, localizada na linha mediana dorsal define o eixo primitivo do embrião, (define a região dorsal e ventral, o lado direito e esquerdo e a região anterior e posterior), também atua na embriogênese do esqueleto axial e representa o elemento estrutural essencial para a locomoção, fornecendo suporte e inserção para os músculos nos cordados ancestrais durante toda a vida.
O que é a notocorda?
A notocorda é um bastão de aspecto cartilaginoso e estruturalmente flexível que se encontra posicionado ao longo do eixo anteroposterior, na região dorsal do corpo dos cordados. É formada por células de origem mesodérmica. A notocorda pode ser dividida em três regiões: uma região interna, medular é formada principalmente por células ricas em fluído citoplasmático onde há abundância de glicoproteínas, e duas bainhas uma interna fibrosa (bainha fibrosa) e uma externa constituída por fibras elásticas formadas de colágeno (bainha elástica). Ao dobrar-se ela volta ao seu estado natural sem gasto de energia, fornecendo um suporte para inserção de músculos atuando no deslocamento nos taxons basais dos cordados.
Origem dos cordados
Existem inúmeras hipóteses para explicar a origem dos cordados, mas temos salientar que virtualmente todos grupos de invertebrados já foram sugeridos como grupos irmãos dos Chordata, inclusive protozoários! Na primeira filogenia conhecida que inclui os vertebrados, Lamarck (1809) evidencia anelídeos e moluscos como grupos irmãos de vertebrados.
Cabe ressaltar que qualquer grupo de invertebrados que escolhamos, não pode ser convertido diretamente da sua forma moderna para um cordado sem uma reorganização drástica. Consultamos os grupos atuais para buscar pistas sobre seus ancestrais. Atualmente são usadas técnicas moleculares para reconstrução filogenética, mas mesmo estas variam de acordo com o método e são também consideradas hipóteses dinâmicas, puramente descritivas. Isto é, não informam sobre como e nem o porque as mudanças ocorreram, o que seria interessante, visto que as hipóteses devem se basear em novidades/mudanças morfológicas com valor adaptativo.
I) Hipótese Annelida e/ou Artropoda
A favor (Saint-Hilaire 1822 (lagostas)
Gaskell & Patten 1912 (ambos grupos);
contras: Cuvier (1830): Hipótese da inversão
Pode-se notar que esta hipótese se baseia em semelhanças análogas e não homólogas. Lovtrup (1977) coloca novamente artrópodes como grupo irmão dos Chordata, ignora embriologia (artrópodes são protostômios), morfologia e o registro fóssil. Baseia-se em fisiologia, química e histologia. Para esse autor, estes caracteres são mais conservados (estáveis) do que a morfologia; assim, se dois animais possuíssem mais substâncias químicas em comum, mais relacionados estariam. Mas, sabemos hoje que a fisiologia dos animais é muito plástica e evolui mais rápido que a morfologia. Dessa forma essa hipótese tem sido desconsiderada, não havendo dados que a corroborem.
II) Ambulacraria/Echinodermata
(Garstang c. 1900) chamada de hipótese Auriculária (nome da larva dos pepinos-do-mar).
À primeira vista parecia forçado que um ancestral como uma estrela ou pepino do mar seriam relacionados aos cordados (devido a presença de pés tubulares, placas de carbonato de cálcio, simetria pentarradial secundária), entretanto, as larvas são algo similares com larvas de um cordado (simetria bilateral, e posições das partes internas). Faixa ciliada circum-oral deu origem ao cordão nervoso; a faixa ciliada ad-oral deu origem ao endóstilo. Da larva diplêurula (ancestral hipotético), teriam surgido também a notocorda e a musculatura segmentar (os miômeros). O sistema nervoso rudimentar serviria basicamente para controlar a natação. Natação é uma pré-adaptação ao aumento do corpo (relação superfície/volume, os cílios não dariam conta de uma natação eficiente, pois o animal teria o mesmo número de cílios, ou os cílios teriam que fazer mais força para mover o animal).
Os cílios ad-orais (não ajudariam na dieta de um animal com maior tamanho corporal, livre natante que se desloca com a boca aberta) transformados em endóstilo teriam maior valor adaptativo.
No entanto, essa hipótese se baseia em outra hipótese, para explicar a origem dos vertebrados. A saber, que em dado momento as larvas de Urocodados (ascídias), considerado até então como grupo irmão de Chordata, sofreram evolução por pedomorfose (por pedogênese provavelmente) e atingiram a maturidade sexual como larvas livre-natantes e não como organismos sésseis (como a maioria das Ascídias). Essa possibilidade enfrenta argumentação contrária por parte de alguns, pois a fase larval tende a ser breve para evitar uma intensa pressão de predação.
(Pedomorfose: é a retenção de características juvenis (ou das formas larvais) nos adultos. Pode ocorrer por diferentes processos como neotenia (crescimento somático retardado) ou pedogênese (desenvolvimento sexual acelerado).
Apesar disto, através do estudo da embriologia constatou-se que são notáveis as semelhanças entre as larvas. Uma boa prática seria comparar as larvas de anfioxos (Cephalochordata) com larvas de Myxini (feiticeiras) e Petromyzon (Ptromyzontida). Os estudos do desenvolvimento embrionário também forneceram fortes evidencias para o agrupamento dos vertebrados como demonstrado nos trabalhos de Ernst Haeckel (1874). Inicialmente Garstang considerava os Hemicordata como intermediários entre Equinodermos e Cordados. Todavia, atualmente os Hemicordados são considerados grupo irmão de equinodermos e o seu respectivo ancestral deveria ser comum aos cordados. Isto na verdade só corrobora com sua hipótese, torando-a mais simples, visto que necessita de um passo a menos de explicação.
III) Hipótese dos hemicordados (Bateson, 1886; Gogdwin, 1917)
Semelhanças: ambos são deuterostômios, ambos apresentam simetria bilateral, fendas na faringe. Entretanto, os hemicordados apresentam Estomocorda (estrutura similar à notocorda) outrora chamada de hemicorda.
Os hemicordados não apresentam tubo neural oco dorsal, e sim uma rede difusa por todo o corpo, inclusive ventralmente. Para complicar, os hemicordados apresentam muitos genes Hox homólogos aos dos vertebrados, codificando as mesmas partes do corpo.
ESTRUTURA DE UM CORDADO GENÉRICO


CARACTERÍSTICAS SINAPOMÓRFICAS DO NOSSO FILO
Sinapomorfias do filo dos cordados
Os cordados apresentam em algum estágio de seu desenvolvimento as seguintes características sinapomórficas:

Notocorda
Tubo neural oco dorsal
Fendas faringeanas
Endóstilo
Cauda pós-anal
Outras características compartilhadas
Coração ventral
Epitélio de revestimento
Miômeros em forma de W
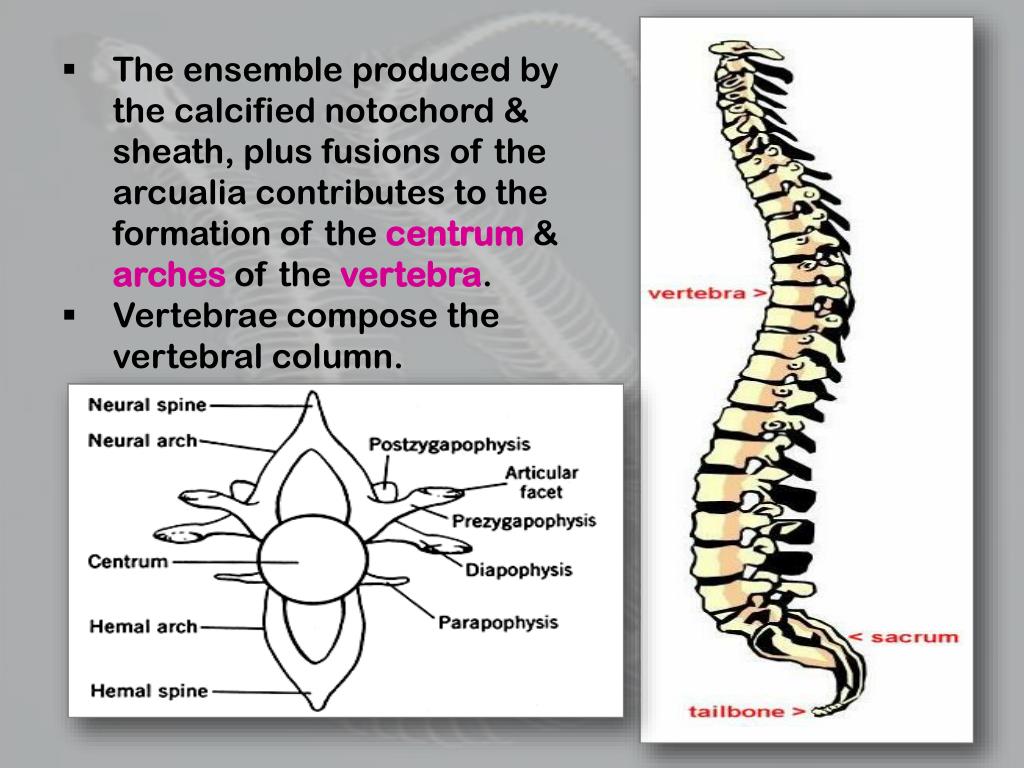
Notocorda
A notocorda é uma estrutura flexível em forma de bastão derivada da mesoderme. Essa estrutura surge logo no início do desenvolvimento embrionário, na gastrulação e ao se formar na região mediana do corpo determina o eixo ântero-posterior do corpo. sua função primordial é fornecer sustentação para o embrião, induzir a formação de outras características essenciais como por ex. o tubo neural oco dorsal. Ela persiste por toda vida e provê suporte para a fixação de músculos nos protocordados (urocordados e cefalocordados) e nos peixes ágnatos (sem mandíbulas). Nos demais grupos, a notocorda está presente somente na fase embrionária, sendo substituída pelas vértebras.
É também chamada de corda dorsal, a notocorda, é uma estrutura mesodérmica de tecido conjuntivo que se inicia no processo notocordal como um tubo oco e posteriormente torna-se maciço. Seu interior é constituído por células grande com vacúolos cheios de liquido, circundadas por uma camada de células semelhantes a uma epitélio. Protegendo toda essa estrutura existem duas bainhas, uma interna chamada de bainha fibrosa e outra externa, a bainha elástica.
A notocorda está presente em pelo menos uma fases do desenvolvimento dos cordados. Sua primeira função é a de sustentação do embrião. Em alguns táxons do filo ela é a única estrutura de sustentação ao longo de toda a vida, em outros ela desaparece ao longo do desenvolvimento (induzindo a formação das vértebras. Nos vertebrados, a notocorda, após o desenvolvimento embrionário, é substituída pela coluna vertebral e irá formar o núcleo pulposo dos discos intervertebrais.
Tubo neural oco dorsal
Estrutura tubular oca, de origem ectodérmica (neuroectoderme) cuja formação é induzida pela notocorda, e localizada dorsalmente à notocorda. Esse tubo nervoso dorsal oco dará origem ao sistema nervoso central e periférico. Na região anterior originará o tronco cerebral, cerebelo e cérebro e a região logo posterior o tronco cerebral constituirá a medula espinhal. Fendas faringianas
Também chamadas de fendas branquiais, ou fendas faríngeas, fenestras branquiais. São aberturas localizadas na faringe que possuem as funções de nutrição (por filtração) e secundariamente respiração, i.e., filtração da água que entra pela boca e assim capturar os alimentos da água e trocas gasosas.
Nos animais aquáticos (exceto amfíbios, répteis, aves, e mamíferos), essas fendas persistem por toda a vida, já nos animais terrestres, essas fendas se mantém apenas na fase embrionária, transformando-se em mandíbula, parte do pescoço e artéria aorta.
Endóstilo
Endóstilo está localizado abaixo das fendas faríngeas e secreta muco que ajuda no acumulo de muitas partículas que vem das fendas. O endóstilo vai dar origem a glândula tireóide. Endóstilo é um sulco ciliado da faringe dos Tunicados, Anfioxos e larvas de
Lampreias, nos adultos de lampreias se metamorfoseia na tireoide (glândula
endócrina responsável por regular o metabolismo dos animais, associado ao
metabolismo do iodo), que é homóloga nos demais Vertebrata, dotado de
cílios e de células secretoras de muco para capturar alimento durante a
filtragem.

Cauda pós-anal
Após a região do ânus, os cordados possuem um prolongamento do corpo sustentado pela notocorda. Nesta estrutura, assim como no corpo estão presente músculos segmentares. A cauda pós-anal confere capacidade de locomoção em meio aquático, uma característica altamente adaptativa. Alguns cordados terrestres mantém a cauda funcional por toda a vida; nestes a cauda funciona como reguladora do equilíbrio quando da perseguição das presas, enquanto outros, como os humanos, a cauda se reduz e suas vértebras se fundem formando o Coccis, estrutura vestigial da cauda.
Outras características compartilhadas
Coração ventral
Vasos pulsantes ventrais, envia sangue oxigenado para o dorso pelos arcos faríngeos e dai para a região posterior por um vaso dorsal, o coração verdadeiro, com câmaras só surge nos Craniata.
Miômeros em forma de V ou W
Os cordados apresentam uma inovação no que se refere à organização dos músculos. Estes estão organizados em séries em forma de V (são músculos segmentares ou miômeros) que são responsáveis pelos movimentos de natação e escavação e são separados uns dos outros por uma membrana de tecido conjuntivo, chamada miossepto.
Epitélio de revestimento
Epitélio especializado nos vertebrados como escamas, placas ósseas, penas e pelos.
localização estrutura e funções da notocorda

Funções da notocorda


Estrutura organização e funções da notocorda
Nos vertebrados a notocorda surge a partir do organizador dorsal (região que pode formar um segundo eixo embrionário). Nesses organismos, o organizador dorsal aparece já durante o período de gastrulação, quando estão se formando os folhetos embrionários. Essa estrutura desempenhará um importante papel de regulador de movimentos celulares na ectoderme e na mesoderme, com a finalidade de continuar o desenvolvimento da notocorda.
Função da notocorda
A notocorda é a estrutura definidora dos cordados e tem papéis essenciais no desenvolvimento dos vertebrados. Ele serve como uma fonte de sinais da linha mediana que padronizam os tecidos circundantes e como um importante elemento esquelético do embrião em desenvolvimento.
Funções principais da notocorda
define o eixo do embrião
serve como base para formação do esqueleto axial
Sinalização para formação dos tecidos do embrião
dirige a formação do tubo neural oco dorsal
dirige a formação das vértebras
ou
A notocorda define o eixo primitivo do embrião.
serve de base para a formação dos ossos da coluna vertebral e da cabeça (esqueleto axial).
indica o local dos futuros corpos vertebrais (dirige a formação das vértebras).
A linha primitiva estabelece o plano básico do corpo nos vertebrados: eixo central (linha média), simetria bilateral (lado ED), região ventral e dorsal e extremidades cefálica e caudal.
O nó primitivo (Nó de Hensen) define a direção cefálica.
Em vertebrados superiores, a notocorda existe temporariamente e tem pelo menos duas funções importantes. Primeiro, a notocorda é posicionada centralmente no embrião em relação aos eixos dorsal-ventral (DV) e esquerda-direita (ED), definindo o eixo da simetria bilateral.
Produz fatores secretados (proteínas) que sinalizam para todos os tecidos circundantes, fornecendo informações de posição e destino. Neste papel, notocorda é importante para especificar destinos ventrais no sistema nervoso central, controlar aspectos da assimetria ED (esquerda/direita), induzir destinos pancreáticos, controlar a identidade arterial versus venosa dos principais vasos sanguíneos axiais e especificar uma variedade de tipos de células na formação dos somitos.
Em segundo lugar, a notocorda desempenha um papel estrutural importante. Como tecido, está mais intimamente relacionado à cartilagem e provavelmente representa uma forma primitiva de cartilagem. Conseqüentemente, a notocorda serve como esqueleto axial do embrião até que outros elementos, como as vértebras, se formem. Em alguns clados de vertebrados, como os agnatha (lampreias), e em peixes primitivos, como o esturjão, a notocorda persiste por toda a vida. Em vertebrados superiores, no entanto, a notocorda torna-se ossificada em regiões de formação de vértebras e contribui para o centro dos discos intervertebrais em uma estrutura chamada núcleo pulposo.

Notocorda dirige a formação do tubo neural

Localização dos somitos da notocorda e do tubo neural

Formação do tubo neural oco dorsal

Formação do tubo neural oco dorsal

Eletromicrografia de varredura de um corte transversal mostarndo os somitos resultantes da organização do mesoderma paraxial bem como o mesoderma intermediário (Mi) e o mesoderma lateral (Ml). TNOD: tubo neural oco dorsal, N: notocorda.




VIDEOS FORTEMENTE RECOMENTADOS
https://youtu.be/hHkmyXLErn4
Formação da notocorda em humanos
https://youtu.be/QVLqxm2GAIg
Gastrulação e formação da notocorda
NEURULAÇÃO
A neurulação acontece quando notocorda envia um sinal químico ao tecido que a recobre para que se torne mais espesso, formando a placa neural. Esta invagina-se originando, assim, o sulco neural. As pregas dentro dele funde-se e fecham para formar o tubo neural. Parte do tecido das pregas originará a crista neural, futuro sistema nervoso periférico.
São processos envolvidos na formação da placa neural e pregas neurais e fechamentos destas pregas para formar o tubo neural.
A neuroectoderma forma o sistema nervoso,
formação do tubo neural oco dorsal,
diferenciação das vesículas cefálicas e da medula espinhal, crista neural,
somitogênese: o mesoderma paraxial e seus derivados.


A notocorda contribui para todos esses fenômenos.
A notocorda é uma estrutura embrionária comum a todos os organismos do filo Chordata, fornecendo pistas mecânicas e de sinalização química para o embrião em desenvolvimento. Em vertebrados, a notocorda surge do organizador dorsal e é crítica para o desenvolvimento adequado dos vertebrados. Essa estrutura evolutiva conservada, localizada na linha mediana dorsal define o eixo primitivo do embrião, atua na embriogênese do esqueleto axial e representa o elemento estrutural essencial para a locomoção, fornecendo suporte e inserção para os músculos nos cordados ancestrais.

Formação do sistema nervoso

Somitogênese

Filo Chordata
Existem cerca de 45 mil espécies de cordados conhecidas, distribuídas em três subfilos: Urochordata (Urocordados), Cephalochordata (Cefalocordados) e Craniata ou Vertebrata.
Os urocordados e cefalocordados não possuem crânio nem coluna vertebral, são invertebrados. Provavelmente, são os cordados mais ancestrais e constituem os Protocordados.
Os craniatas são todos os vertebrados e representam cerca de 98% das espécies deste filo.
CARACTERÍSTICAS GERAIS DO FILO CHORDATA
Sinapomorfias do filo dos cordados:
Notocorda
Tubo neural oco dorsal
Fendas faringeanas
Endóstilo
Cauda pós-anal

Larva de salpa (Ascídia) Urocordado

Cefalocordado atual, anfioxo (Branchiostoma lanceolatum)

Cordado hipotético e suas sinapomorfias
São organismos Triblásticos, Deuterostômios, Simetria bilateral, Enterocelomados, Metamerizados.
Metaspriggina e Pikaia
O conhecimento da evolução inicial dos cordados e dos peixes depende em grande parte do material de corpo mole do período Cambriano Inferior (Série 2) do Sul da China. Devido à raridade de algumas dessas formas e à falta geral de material comparativo de outros depósitos no mundo, as interpretações de várias características permanecem controversas, assim como suas relações mais amplas entre os primeiros vertebrados sem mandíbula não esqueletizados pós-cambrianos.
Numa redescrição de Metaspriggina com base em novo material de Burgess Shale e material excepcionalmente preservado coletado perto de Marble Canyon, British Columbia, e três outros depósitos cambrianos semelhante aos de Burgess Shale de Laurentia.

Metaspriggina sp um cordado ancestral do cambriano
Este peixe primitivo exibe características inequívocas de um vertebrado: uma notocorda, um par de olhos proeminentes do tipo câmera (existem três tipos de olhos na natureza: olhos compostos, olhos tipo espelhos e olhos do tipo câmera), sacos nasais emparelhados, possível crânio e arcualia, miômeros em forma de W e uma cauda pós-anal, todas essas características estão presentes nos cordados atuais.


Formação e evolução da notocorda

Uma característica marcante é a área branquial com uma série de barras bipartidas. Com exceção da barra mais anterior, que parece ser um pouco mais espessa, cada uma está associada a brânquias localizadas externamente, possivelmente alojadas em bolsas. A análise filogenética coloca Metaspriggina como um vertebrado basal, aparentemente próximo aos táxons de Chengjiang Haikouichthys e Myllokunmingia, demonstrando também que este grupo ancestral de peixes era cosmopolita durante os tempos do Cambriano Médio-Inferior (Série 2-3). No entanto, o arranjo da região branquial em Metaspriggina tem implicações mais amplas para reconstruir a morfologia do vertebrado primitivo. Cada barra bipartida é identificada como sendo respectivamente equivalente a um epibranchial e ceratobranchial. Essa configuração sugere que um arranjo bipartido é primitivo e reforça a visão de que a cesta branquial de lampreias provavelmente é derivada. Outras características do Metaspriggina, incluindo a posição externa das brânquias e a possível ausência de uma brânquia oposta à barra mais anterior mais robusta, são características dos gnatostomados (peixes com mandíbulas) e, portanto, podem ser primitivas dentro dos vertebrados.


Adaptado de Land, 2005.
Laurentia
Laurentia ou Cráton da América do Norte é um grande cráton continental que forma o antigo núcleo geológico da América do Norte. Muitas vezes no passado, Laurentia foi um continente separado, todavia hoje está formando parte América do Norte, embora originalmente também incluísse as áreas cratônicas da Groenlândia e também a parte noroeste da Escócia, conhecida como "terranes" das Hébridas (terranes: é um fragmento de material da crosta formado ou separado de uma placa tectônica e acrescido ou "suturado" à crosta que está em outra placa). Em outras épocas do passado, Laurentia fez parte de continentes e supercontinentes maiores e elo próprio consiste em muitos "terranes" menores montados em uma rede de cinturões orogênicos do Proterozóico Inferior (proterozócio é um éon geológico que abrange o tempo desde o aparecimento do oxigênio na atmosfera da Terra até um pouco antes da proliferação de vida complexa, como trilobitas ou corais, na Terra). Pequenos microcontinentes e ilhas oceânicas colidiram e se suturaram na Laurentia, que sempre crescia, e juntos formaram o cráton pré-cambriano estável visto hoje. O cráton é nomeado pelo o Escudo Laurentiano, através das Montanhas Laurentian, que recebeu seu nome do Rio São Lourenço, em homenagem a Lourenço de Roma.

Laurentia, também chamado de crato norte-americano
(Fonte: Wikipedia)
Cientistas chineses e britânicos descobriram em rochas do período Cambriano (545 milhões a 490 milhões de anos atrás) os mais antigos vestígios de animais vertebrados de que se tem notícia. São bisavós de peixes, ainda desprovidos de mandíbulas, que tiveram seus corpos quase milagrosamente preservados. A descoberta recua entre 20 milhões e 50 milhões de anos a datação de fósseis de vertebrados (animais dotados de coluna vertebral, como peixes, aves e mamíferos).
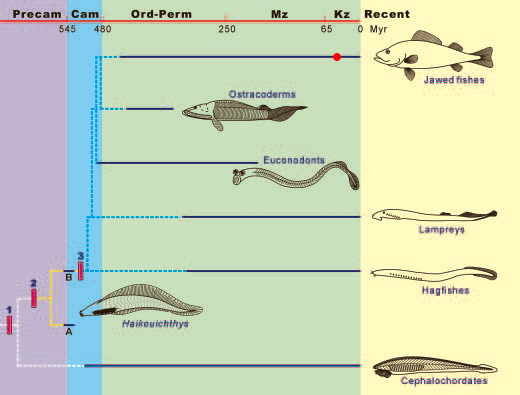
Origem dos peixes no cambriano e sua diversificação
Entre as implicações científicas do achado está um reforço para a tese de que ossos são uma invenção relativamente recente da natureza. Os nomes atribuídos às duas espécies descobertas são: Haikouichthys ercaicunensis e Myllokunmingia fengjiaoa.


Haikouichthys ercaicunensis

Haikouichthys ercaicunensis

O vertebrado agnatha do Cambriano Inferior Haikouichthys ercaicunensis Luo, Hu & Shu gen. et sp. nov. de Haikou, Yunnan. Specimen HZ-f-12-127. a, Amostra inteira, anterior à esquerda; a região mais posterior parece desaparecer em sedimento, possivelmente representando a decomposição do corpo; as tentativas de escavar esta área não tiveram sucesso. Barra de escala equivalente a 5 mm. b, Detalhe da parte anterior para mostrar barras de brânquia (supostas), possíveis elementos do endoesqueleto craniano e área pericárdica; barra de escala equivalente a 5 mm. c, Desenho em câmera-lúcida de espécime para mostrar a interpretação. Os números 1 a 6 indicam as unidades da cesta branquial que são identificadas com alguma segurança; ? A–? C referem-se a identificações menos seguras. São indicadas duas áreas possíveis que representam a cavidade pericárdica. Para a parte anterior de C, uma área triangular com manchas de mineralização diagenética é uma possibilidade; uma região mais fraca na parte posterior é o local alternativo.

Tamanho do Haikouichthys ercaicunensis quando comparado com um ser humano


Análise filogenética dos primeiros vertebrados, incluindo Myllokunmingia gen. nov. e Haikouichthys gen. nov. a, Árvore de consenso estrito das seis árvores mais parcimoniosas. b, uma das seis árvores mais parcimoniosas; os outros cinco devem apresentar diferenças mínimas.
Os paleontólogos que os descobriram relatam que não encontram sinal algum de mineralização em seus tecidos, ou seja, eles não tinham ossos. Isso ajuda a explicar por que são tão raros os fósseis desses ancestrais de todos os vertebrados. Tecidos ósseos são mais propícios à fossilização, (a fossilização não passa da substituição de substâncias orgânicas por minerais, e para que isso aconteça uma estrutura rígida como os ossos que levam mais tempo para serem atacados por microrganismos e condições especiais de tafonomia são essenciais). Estruturas mais moles, como cartilagens, couro, pele e órgãos internos, muito raramente produzem fósseis. Da época do Cambriano só se conhecem dois sítios em que esses tecidos moles foram preservados: a pedreira de Burgess Shale, no Canadá, e Chengjiang, na China. Desses dois locais provém a coleção de fósseis de uma fauna espetacular, chamada de fauna Ediacara. O repentino surgimento de tantos e tão diferentes "bauplans" de metazoários multicelulares no registro geológico ficou conhecido na literatura como explosão Cambriana. Até agora, uma ausência notável era percebida nessa época: ossos e vértebras da coluna. Havia indícios de cordados, o filo que inclui peixes, anfíbios, répteis, aves e mamíferos, todavia bastante controverso.
Os vestígios encontrados por Simon Conway Morris, da Universidade de Cambridge (Reino Unido), e seus colegas chineses são inequivocamente de vertebrados. Crânios, branquias, nadadeiras, tudo parece estar lá. "Os dois novos fósseis (...) da localidade de Chengjiang são os mais convincentes vertebrados do Cambriano jamais encontrados", relata o paleontólogo Philippe Janvier, que não participou da descoberta. Conway Morris é um grande nome da paleontologia, e não só para o público especializado. Ele liderou a escavação em Burgess Shale, a primeira formação a revelar a estranha fauna Ediacara.
Stephen Jay Gould defendia em seu livro "Vida maravilhosa" a idéia de que a explosão cambriana foi um gigantesco experimento da natureza com as mais bizarras arquiteturas corporais. Somente umas poucas teriam dado certo, entre elas estava a que originaria os vertebrados. Gould, enfatizou a grande participação do acaso nesse processo, alem da seleção natural atuando sobre os fenótipos ao londo de molhoes de anos. Vale ressaltar que Conway Morris lançou uma obra sobre a fauna de Burgess shale, "The Crucible of Creation - The Burgess Shale and the Rise of Animals". (Modificado da Folha de São Paulo)
A espécie Metaspriggina walcotti, revelou-se como um dos nossos ancestrais, um tipo de tetravô de todos os vertebrados. Nos oceanos primitivos do período Cambriano onde surgiram os primeiros animais, há cerca de 500 milhões de anos. Só dois espécimes incompletos eram conhecidos, e agora o líder do campo, Simon Conway-Morris, descobriu uma centena deses animais no Canadá, entre os quais muitos estão em bom estado de conservação.

Diagrama de Metaspriggina, o cordado mais basal. Lectótipo –USNM198612 e ex-holótipo 198611 no Museu Nacional de História Natural, Smithsonian Institution, Washington, DC, EUA. Este “verme” de natação livre é próximo aos ancestrais dos vertebrados, incluindo humanos, em última análise.
As primeiras bactérias apareceram na Terra há aproximadamente 3,5 bilhões de anos, não muito depois do surgimento do planeta e do Sistema Solar, o que indica que a vida surge muito rápido quando ha condições propícias e evolui também rapidamente atraves da seleção natural. Durante 75% do tempo transcorrido desde então, contudo, não houve qualquer aumento de complexidade. Os primeiros animais só surgiram há cerca de 550 milhões de anos, em um acontecimento tão espetacular para a escala geológica que, na literatura científica, recebe o nome de explosão cambriana. E o fóssil de Metaspriggina apareceu nessa época.
A anatomia comparada clássica, criada pelo naturalista francês do século XVIII George Cuvier, considerava tão insuperáveis as diferenças do plano corporal existente entre um molusco (mexilhão), um artrópode (mosca) e um vertebrado (peixe), que imaginou que esses planos corporais eram "obras independentes do um criador", surgidas do laboratório divino como ideias essencialmente distintas, como se procedessem de diferentes projetos ou planos. No tempo de Darwin, entretanto, já estava claro que todos esses planos de design corporal (baupläne ou bauplan) surgiram simultaneamente na explosão cambriana. Todavia, Darwin sempre considerou essa origem brusca de todos planos corporais como um empecilho à sua teoria da evolução através da seleção natural.
Hoje sabemos que não, e o trabalho de Simon Conway-Morris, da Universidade de Cambridge, mostra isso. Inspirado pela leitura dos artigos de Conway-Morris que trabalha há mais de 30 anos, com os fósseis da pedreira de Burgess Shale, Stephen Jay Gould sua obra mais lida e influente: "It’s a wonderful life", ou A vida maravilhosa, um dos grandes best-sellers científicos da segunda metade do século XX.
Gould imaginou, como também o próprio Conway-Morris, que a explosão cambriana revelava a imensa capacidade de inovação da vida, ao criar vinte planos corporais radicalmente distintos dos quais sobreviveram apenas cinco, entre eles está o bauplane cordado. O fóssil de Metaspriggina walcotti é a última de uma série de evidências que, na verdade, revelam que os aparentemente surreais planos de desenho da explosão cambriana não são mais que versões iniciais dos animais atuais, como é o Metaspriggina em relação ao bauplan dos vertebrados. Ou seja há uma continuidade entre as formas o que pode ser explicado pela seleção natural atuando ao longo de um grande lapso de tempo.
E que, de fato, os moluscos os artrópodes e vertebrados apresentam uma origem comum situada pouco antes do cambriano, e que não somos mais do que variações de uma arquitetura elementar de grande simplicidade. E as variações que são encontradas no registro fóssil são respostas a diferentes pressões ambientais sobretudo aquela apresentada pela quantidade de oxigênio existente na Terra naquela época.

Cordados ancestrais
Espécies de cordados ancestrais: (A) Didazoon, um vetulicoliano (um grupo possivelmente relacionado a cordados) do cambriano inferior (B) Yunnanozoon, um hemicordato ou cordado do cambriano inicial (C) Haikouella, um possível chordado do cambriano inferior (D) Cathaymyrus, um cefalocordado do cambriano (E) Pikaia, um cefalocordado do cambriano (F) Branchiostoma, um cefalocordado (G) Metaspriggina, um cordado do cambriano médio (H) Myllokunming, um possível vertebrado basal do cambriano inferior (I) Haikouichthys, um vertebrado basal do cambriano (J) um Conodonte, um cordado que viveu do cambriano ao triássico superior (K) Pharyngolepis, um vertebrado agnato do siluriano (L) Jamoytius, um agnatho vertebrado do siluriano (M) peixe-bruxa, agnatho chordata (N) uma lampreia, agnatho vertebrado atual. Os organismos não estão em escala.
Triblásticos, Deuterostômios, Simetria bilateral, Enterocelomados, Metamerizados

SINAPOMORFIAS DOS CORDADOS









PROTOCORDADOS
O termo Protocordados refere-se a um grupo possivelmente parafilético (por excluir os vertebrados) de animais invertebrados que apresentam clivagem radial do zigoto.
São deuterostômios marinhos que possuem fendas faríngeas.
O termo não consiste de um nome científico e válido, mas sim é uma denominação de um agrupamento de organismos mais simples e mais próximos aos Chordata.
A reunião de organismos tão diversos em um grupo próximo aos vertebrados baseia-se principalmente em aspectos do desenvolvimento embrionário.
Na fase de nêurula, todos os protocordados ou cordados verdadeiros (vertebrados) exibem o mesmo padrão de organização do corpo, sendo possível identificar as quatro estruturas que caracterizam o grupo: uma notocorda, um cordão nervoso oco dorsal, fendas brânquias ou faríngeas e uma cauda pós-anal.
A notocorda é um bastão flexível de células, que se estende por, praticamente, toda a região dorsal do corpo do animal, localizando-se acima do sistema digestório e abaixo do sistema nervoso.
Ela serve como um ponto de apoio para os músculos auxiliando nos movimentos desses animais.
Nos protocordados, a notocorda, uma estrutura de sustentação, formada por tecido conjuntivo, não é substituída por tecido ósseo; portanto, não forma uma coluna vertebral, como ocorre com os vertebrados.
Cordão nervoso oco dorsal
A maioria dos filos estudados até agora possui um sistema nervoso que se situa na região ventral e é formado por um cordão duplo e maciço de células. Já entre os protocordados e os cordados, esse é um cordão dorsal, simples e oco.
Fendas faríngeas
As fendas branquiais ou faríngeas, pares, situam-se na região da faringe e exercem função respiratória. Entretanto, nos vertebrados terrestres, são substituídas por pulmões, durante o desenvolvimento embrionário.
Cauda pós-anal
A cauda pós-anal, muscular, esta situada na região posterior ao ânus. Agora que você já conhece, intimamente, as características básicas dos protocordados e dos cordados, deve estar se questionando como esses animais surgiram.
Endóstilo
Sulco ciliado com função de captura de alimento.
Como surgiram os protocordados
Algumas teorias foram elaboradas a fim de explicar o parentesco evolutivo dos protocordados. Uma das primeiras teorias, que foi proposta no século XIX, supunha que os protocordados teriam se originado a partir de anelídeos marinhos, devido a sua simetria bilateral, segmentação do corpo e tubo nervoso longitudinal ventral.
Posteriormente, foi defendida a teoria que relacionava os cordados aos equinodermos, por apresentarem endoesqueleto, serem deuterostômios, possuírem simetria bilateral e apresentarem, bioquimicamente, algumas proteínas musculares e alguns amebócitos sanguíneos. Contra essa teoria, temos a simetria radial secundária e o sistema ambulacral.
Sobre a simetria radial trata-se de um argumento frágil, pois a simetria inicial dos equinodermos é a simetria bilateral.
Uma terceira teoria defende que os cordados e os equinodermos têm a mesma origem evolutiva, ou seja, um ancestral comum. (Barbosa e Noveli, 2009; Martho, 2019).
Os dois subfilos de animais deuterostômios que apresentam notocorda em pelo menos algum período do desenvolvimento embrionário, e que são reunidos como protocordados, são os Urochordata ou Tunicata, subfilo que reúne não só as ascídias como também as salpas, e os Cephalochordata, subfilo representado pelos anfioxos.
Devemos lembrar que no passado, os hemicordados eram classificados dentro dos protocordados, mas a análise de sua provável notocorda os separa desse agrupamento.
Os urocordados são considerados mais basais, enquanto os cefalocordados são considerados grupo-irmão dos vertebrados, por partilharem a presença de miomeria, ou seja, a repetição de pacotes musculares ao longo do corpo, e pelo fato de a notocorda estender-se até a região da cabeça.
Hipóteses sobre relação entre os Chordata
Os Chordata podem ser divididos em 3 grupos:
Urochordata (Tunicados / Ascídias),
Cephalochordata (Anfioxo) e
Craniata (Vertebrados).
Entre 1928 até hoje diversas árvores foram propostas com todos arranjos possíveis. Mas, baseado em uma análise molecular complexa em 2006 os Tunicados são colocados como grupo irmão dos Craniata.
Em 2008 um novo estudo molecular coloca os Cephalocordados como grupo irmão (realmente Cephalochordata tem mais semelhanças, inclusive morfológicas).
Em 2009 voltam a colocar os Tunicados como irmão. Assim, vejam que a situação ainda não está definida.
Em uma análise recente (2008) sobre filogenia funcional (levando-se em consideração não só a morfologia, mas também a função das estruturas, algo que tem se mostrado mais preciso do que a fenética tradicional), Tunicata e Acrania (=Cephalochordata) são grupos irmãos e seu ancestral comum divergiu de Craniota (Craniata).
Independente de quem é o grupo irmão de quem, é importante conhecermos todos, pois o grupo irmão dos vertebrados nos ajuda a entender a origem dos vertebrados e o grupo que ficar mais basal na filogenia (e os anfioxos estão sendo os mais cotados para assumir essa posição), nos ajuda a compreender a origem dos Chordata.
Além disso, todos sendo cordados devem compartilhar semelhanças (caracteres derivados exclusivos).
São as famosas 5 sinapomorfias:
1) Presença de Notocorda, em ao menos uma fase da vida.
2) Cordão nervoso oco dorsal
3) Cauda pós-anal, em ao menos uma fase da vida
4) Fendas faríngeas, em ao menos uma fase da vida: usada para filtragem de alimentos em não vertebrados e primariamente como órgão respiratório em vertebrados aquáticos (peixes). Mas essa estrutura também é observada em outros deutereostômios (grupo que compreende Chordata), assim, deve ser uma plesiomorfia e não uma sinapomorfia. A função das fenestras branquiais é primariamente para alimentação. Respiração é uma função secundária, portanto uma característica derivada.
5) Endóstilo, sulco ciliado da faringe dos Tunicados, Anfioxos e larvas de Lampreias. Nos adultos de lampreias se metamorfoseia na tireoide (glândula endócrina responsável por regular o metabolismo dos animais, associado ao metabolismo do iodo, que é homologa nos demais Vertebrata, dotado de cílios e de células secretoras de muco para capturar alimento durante a filtragem.
0 comentários:
Postar um comentário