CLASSIFICAÇÃO E EVOLUÇÃO
CAROLUS LINNAEUS E A SISTEMÁTICA

Dryadula phaetusa L. 1758 (Nymphalidae; Heliconiinae)
(Fonte: Steven Erat, 2006)



CAPA DO LIVRO DE LINEU (EDIÇÃO DE 1758)

KARL LINNÉ (1707 - 1778)
As borboletas Heliconius têm duas características ecológicas derivadas únicas que podem ter facilitado a rápida radiação adaptativa: alimentação com pólen e comportamento de acasalamento de pupas (pupal-mating behaviour) (Gilbert, 1972).
As borboletas adultas coletam sistematicamente o pólen das flores, que mastigam na tromba para dissolver os aminoácidos. Isso permite que as lagartas se desenvolvam de forma relativamente rápida (uma vez que não precisam armazenar nutrientes para a produção de óvulos e espermatozoides) e permite que os adultos tenham uma vida útil bastante prolongada, até 8 meses, na natureza.
Heliconius butterflies have two unique, derived ecological traits that may have facilitated rapid adaptive radiation: pollen feeding and pupal-mating behaviour (Gilbert, 1972). Adult butterflies systematically collect pollen from flowers, which they masticate on the proboscis to dissolve out amino acids. This allows caterpillars to develop relatively rapidly (since they do not need to store nutrients for egg and sperm production), and allows adults to have a greatly extended lifespan, up to 8 months – in the wild. (Beltrán, Brower and Jiggins, s/d).
According to Beltrán, et alii, s/d, the pollen-feeding behaviour centres around a group of vines in the Cucurbitaceae, Psiguria and Gurania.
This is a tight plant-pollinator relationship in which the butterflies are major pollinators for the plants and the plants major food resources for adult maintenance and egg production. An amount of 80% of a female's egg production come from amino acids that come from pollen she collects.
Only 20% comes from amino acids acquired by the caterpillars feeding on passion vines. In most butterflies and moths, 100% of eggs derive from the efforts of the larval stage, and eggs are laid in a quick pulse after adult emergence.
In Heliconius, eggs are laid as they are manufactured over the adult's long lifespan (Gilbert, 1972). The butterflies learn the locations of pollen plants and establish home ranges based on pollen foraging routes. It appears that the pollen plants are more significant than larval hosts in determining a female's assessment of the habitat.
Thus, as long as she knows the locations of a network of pollen plants, she will stay in the area, even during periods when new shoots of passion vine hosts are temporarily not available due to weather or defoliation by Heliconius or other competing herbivores. So while most herbivorous insects disperse away when suitable oviposition sites are scarce, Heliconius females are content to stay put as long as the mutualist plant produces pollen (which is year-round). Moreover the pollen promotes a long reproductive life so that females can wait many weeks for the opportunity to resume egg-laying (Ehrlich & Gilbert, 1973).(tolweg)
A second unusual trait found in some Heliconius species is a unique mating behaviour known as pupal-mating. Males of certain species search larval food plants for female pupae. The males then sit on the pupae a day before emergence, and mating occurs the next morning, before the female has completely eclosed (Gilbert, 1976; Deinert et al. 1994). Various kinds of pupal-mating occur scattered across several insect orders (Thornhill & Alcock, 1993); in passion-vine butterflies almost half the Heliconius species (42%) are pupal-maters (Gilbert, 1991, pupal mating clade marked in the cladogram above).
Gilbert (1991) suggested that pupal-mating might play an important role in the radiation of Heliconius as well as in the packing of Heliconius species into local habitats. Pupal-mating might enhance the possibility of intrageneric mimicry because in many cases, mimetic species pairs consist of a pupal-mating and a non pupal-mating species. The strikingly different mating tactics of these groups could allow phenotypically identical species to occupy the same habitats without mate recognition errors. Second, this mating tactic may influence host-plant specialisation, as it has been suggested that pupal-mating species may displace other heliconiines from their hosts by interference competition (Gilbert, 1991). Males of these species sit on, attempt to mate with, and disrupt eclosion of other Heliconius species of both mating types encountered on the host plant. This aggressive behaviour may prevent other heliconiine species from evolving preference for host plants used by pupal-mating species.
Etymology: Heliconius signifies dwellers on Mount Helicon (Turner, 1976) (see each species for more information).
Helicon is a mountain in southern Greece, in Boeotia, regarded in ancient Greece, as the source of poetry and inspiration. From it flowed the fountains of Aganippe and Hippocrene, associated with Muses.
The nine Muses are daughters of Zeus and Mnemosyne, the goddess of memory. The Muses sat near the throne of Zeus, king of the gods, and sang of his greatness and of the origin of the world and its inhabitants and the glorious deeds of the great heroes.(Beltrán, Brower and Jiggins, s/d)
From their name words such as music, museum, mosaic are derived (Muses). The nine muses are:
- Calliope: Epic and heroic poetry and the head muse
- Clio: History
- Erato: Love-poetry
- Euterpe: Music and Lyric poetry
- Melpomene: Tragedy
- Polyhymnia: Sublime hymns or serious sacred songs
- Terpsichore: Dancing and choral song
- Thalia: Comedy and idyllic poetry
- Urania: Astronomy
Linnaeus (1758) named various butterflies (all in the genus Papilio section "Heliconius") after the muses, all of which have longish wings, occur in the neotropics (except two) and participate in mimicry complexes, but most of which are not currently placed in the genus Heliconius and two of which are not even in Nymphalidae. These are:
- Stalachtis calliope (L.), 1758 (Riodinidae)
- Eresia clio (L.), 1758 (Nymphalinae)
- Heliconius erato (L.), 1758 (Heliconiinae, South american)
- Stalachtis euterpe (L.), 1758 (Riodinidae)
- Heliconius melpomene (L.), 1758
- Mechanitis polymnia (L.), 1758 (Ithomiini) (a variant spelling of Polyhymnia)
- Acraea terpsichore (L.), 1758 (Acraeini, African)
- Actinote thalia (L.), 1758 (Acraeini)
- Taenaris urania (L.), 1758 (Amathusiini, Australasian)


Borboletas Heliconius com a probóscide contendo pólen coletado de flores. A dieta da maioria dos lepidópteros são muito limitadas em compostos nitrogenados e acredita-se que a alimentação com pólen aumenta a longevidade e a produção de ovos nas borboletas Heliconius. Imagens © Mathieu Joron.
As borboletas Heliconius são reconhecidos por seus grandes olhos, antenas longas, forma de asa alongada característica, célula discal da asa posterior em forma de lágrima e padrões de cores distintos. As plantas hospedeiras são todas Passifloreae, e há alguma associação filogenética entre grupos de espécies de Passiflora e as espécies de Heliconius que se alimentam delas (Benson et al., 1976; Brower, 1997) (ver cada espécie para mais detalhes).
Discussão das relações filogenéticas
Para uma discussão sobre a monofilia do gênero conforme apresentada aqui e as relações entre os gêneros heliconiine, consulte a página Heliconiini.

A sample of the morphological diversity of wing patterns in Heliconius and related genera. Each row represents a phylogenetic clade in the tribe Heliconiini.
© Chris Jiggins and Mathieu Joron

Cladograma para o gênero Heliconius
baseado em três mtDNA e quatro regiões de genes nucleares.
Dentro de Heliconius, as relações apresentadas aqui são baseadas em dados de sequência molecular para três mtDNA e quatro regiões de genes nucleares (Beltran et al. 2007). Há também um clado monofilético com "pupal-mating behaviour" fortemente suportado, sugerindo que o comportamento de acasalamento de pupa evoluiu apenas uma vez na Heliconiinae (ver árvore acima). Dentro de Heliconius, a ausência de um signum na bursa copulatrix feminina é um caráter morfológico que define o grupo com "pupal-mating behaviour" (Penz, 1999).
Distribuição geográfica
Members of the genus are found from the southern United States throughout Central and South America and the West Indies, with the greatest diversity of species in the Amazon Basin (Emsley, 1965; DeVries, 1987). (Beltrán, Brower and Jiggins, s/d)
Heliconius butterflies show a continuum of geographic divergence and speciation; they are unpalatable and exhibit inter- and intraspecific diversification of colour and patterns. Bates’ classic paper (Bates, 1862), reflecting observations during his stay in the Amazon, showed a geographical pattern for the different colour forms: similar between species within any one area of the Amazon basin, but the mimetic colour patterns themselves changed every 100-200 miles. Beside this geographic divergence, closely related species within an area often belonged to mimicry “rings” (groups of unpalatable species, together with some palatable species, that have converged on the same warning colour pattern) (Brower et al., 1964; Mallet and Gilbert, 1995). Bates’ system has all the intermediate stages between local varieties, geographic races, and sympatric species that make it an excellent biological model to study selection, hybridization and gene flow at the species boundary. See maps attached to each species.(Beltrán, Brower and Jiggins, s/d).
Heliconiinae
In the last sixty years seven major studies have addressed the systematics of the passion-vine butterflies or Heliconiini (Michener, 1942; Emsley, 1963, 1965; Brown, 1981; Brower, 1994a; Brower & Egan, 1997; Penz, 1999). These phylogenetic hypotheses are in conflict with one another, in particular with regard to the relationships among the genera Heliconius, Eueides, Neruda and Laparus, leading to disagreement over the origin of the derived behavioural trait, pollen feeding. The most recent attempt to resolve the phylogenetic relationships in the group, using molecular markers, showed that Heliconius is paraphyletic, with Laparus doris and Neruda falling within the genus (Beltran et al., 2007). This analysis was based on molecular sequence data for 3 mtDNA and 4 nuclear gene regions. The results would imply a single origin for pollen feeding but with a loss of the trait in Neruda. However different genes are not congruent in their placement of Neruda, such that statistical analysis implies that the traditional placement of Neruda as a sister group to Heliconius cannot be ruled out. Hence, we here place Neruda as sister to Heliconius, but subsume the monobasic genus Laparus into Heliconius (Heliconius doris).
The most recent morphological analysis of the group (Penz, 1999) differs from the tree presented here in three aspects: (1) the relationships among Podotricha, Dryadula, Dryas and Philaethria; (2) the relationships among Laparus, Neruda, Eueides and Heliconius; and (3) the resolution of basal nodes of the tribal tree. We would therefore caution that the relationships presented here might change given more data – either morphological analysis with more complete species sampling, or molecular data with more fast-evolving nuclear gene regions.
Note that we here consider the ‘passion vine butterflies’ as a tribe, Heliconiini, although others have previously ranked them as a sub-tribe Heliconiina (or "Heliconiiti") (Harvey, 1991; Brower and Egan, 1997; Brower, 2000; Beltrán et al. 2007).
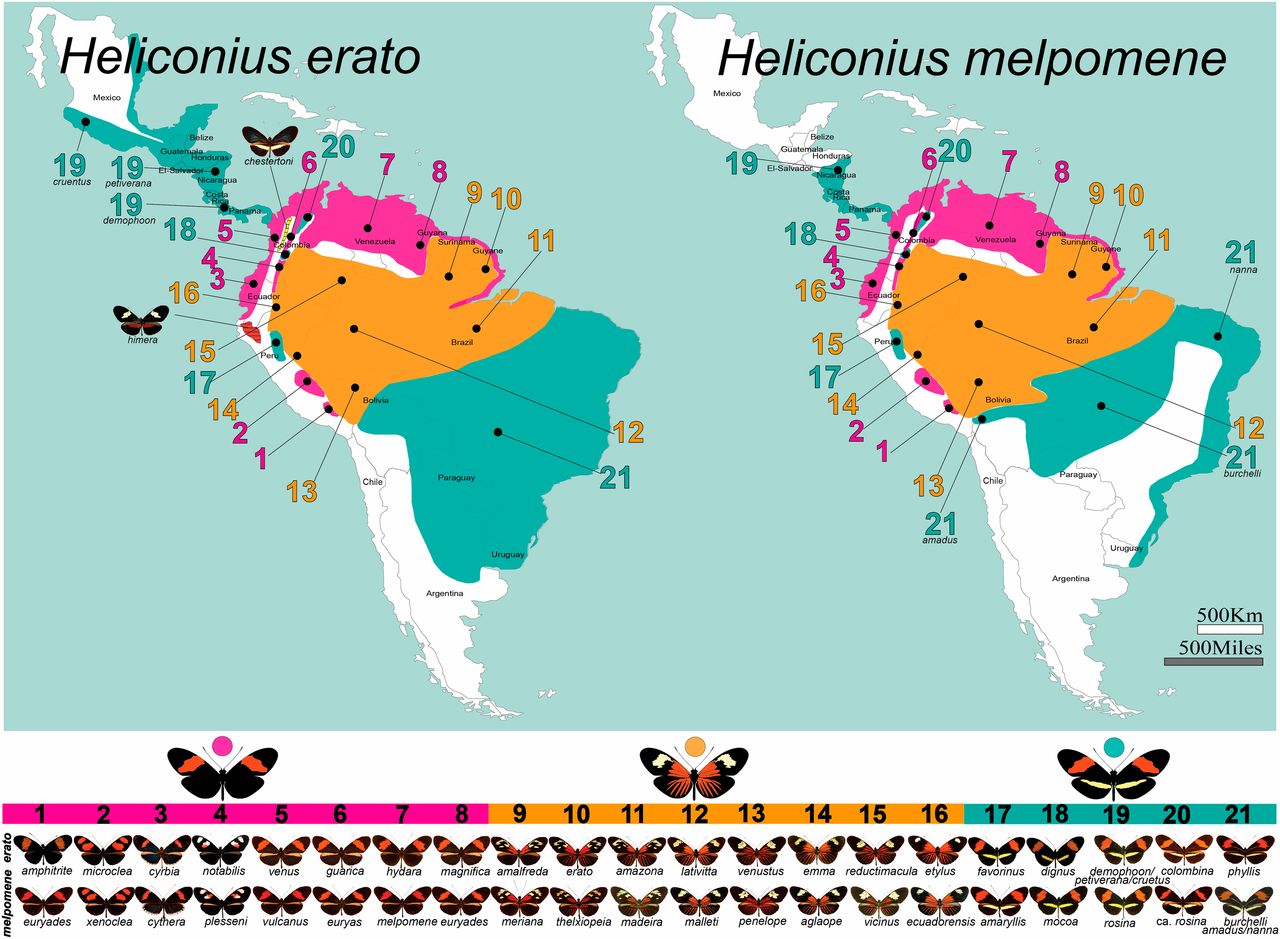
Geographic distributions of the Heliconius erato and H. melpomene convergent radiations. Approximate distributions of the three major phenotypes and locations of many described subspecies, or “races,” of the Müllerian co-mimics H. erato and H. melpomene. Matching wing-color-pattern variation and subspecies names are shown below the maps. Two incipient species that are part of the H. erato radiation, H. himera and H. erato chestertoni, are also presented.

Mapping color-pattern variation in Heliconius: Mendelian segregation, quantitative variation, and genome-wide association. (A) An F2 mapping family from a cross between sister species H. himera and H. erato notabilis shows Mendelian segregation of black (Sd = WntA) and red (D = optix) wing-scale distributions (Papa et al. 2013). (B) Quantitative wing-color variation and linkage group distribution of QTL for black, red, and yellow/white color-pattern variation (stars) in the same mapping family. Stars indicate the presence of at least one locus modulating black (black star), red (red star), or yellow/white (yellow star) pattern variation. (C) Genome-wide association study of wing-color-pattern variation in H. melpomene and H. erato Ecuadorian hybrid zones (Nadeau et al. 2014). The different colors of points represent individual SNPs associated with distinct pattern elements shown in the wing images above. Points above the lines represent significant associations. Red arrows indicate the positions of optix (D locus) and WntA (Sd locus). Blue arrows indicate the positions of putatively undescribed color-pattern loci, many of which are not shared between H. melpomene and H. erato. (Kronforst and Papa, 2015)

Phenotypic diversity across Heliconius butterflies. Phylogenetic relationships among all major Heliconius clades are shown (Kozak et al. 2015). Names and color patterns of representative species and subspecies are depicted, together with behavioral traits that characterize phylogenetic nodes across the genus.

Characterizing the H. numata mimicry supergene. (A) A single Mendelian locus with multiple alleles (the P locus) controls wing-pattern diversity in H. numata. The P locus is positionally homologous to the Yb-Sb-N loci of H. melpomene and the Cr locus of H. erato (Joron et al. 2006b). (B) Fine-mapping and SNP associations narrow the P locus to a 400-kb interval spanning 31 genes and provide evidence of highly reduced recombination (red and blue areas) (Joron et al. 2011). Relative position of the genes (1–7) across the interval that were used to characterize genomic rearrangements is also shown. (C) Allelic variation at the P locus ultimately traces back to an inversion polymorphism with different wing-pattern morphs determined by distinct, nonrecombining haplotypes (Joron et al. 2011). PCR assay of the alternative breakpoints BP0, BP1, and BP2 (left) are perfectly associated with mimicry variation across four distinct morphs in eastern Peru (right).(Kronforst and Papa, 2015).

Phylogenetic relationships between DNA sequences from different Heliconius taxa within the region containing optix (left), compared to the phylogeny of the species based on multiple genes (right). Around optix, taxa cluster according to their colour pattern, not according to the species they belong to.
SISTEMÁTICA
RAMO DA BIOLOGIA QUE PREOCUPA-SE EM INVENTARIAR E DESCREVER AS ENTIDADES VIVAS (ESPÉCIES).
TAXONOMIA
RAMO DA BIOLOGIA QUE PREOCUPA-SE COM A NOMEAÇÃO DE ENTIDADES VIVAS (ESPÉCIES), BEM COMO ESTUDAR SEU GRAU DE RELACIONAMENTO COM OUTRAS ENTIDADES (SERES VIVOS).

Uma das primeiras representações de uma árvore filogenética feita por
Ernst Haeckel (1866)
Ernst Haeckel (1866)
CLADOGÊNESE
Chama-se cladogênese (do grego: Κλάδος = clados= ramo + Γένεσις = gênese= gerar, origem, nascimento) o conjunto de processos que promovem a especiação, i.e., a separação de uma população em duas ou mais e sua subsequente diferenciação genética e morfológica o que leva ao surgimento de novas espécies (equilíbrio pontuado). Ex. no arquipélago Hawaino, onde colonizadores (pássaros e moscas das frutas) viajaram enormes distâncias via correntes marinhas e de vento, chegando aos inúmeros vales de muitas ilhas) se estabelecendo nesses habitats e se diversificando e com o tempo originaram novas espécies (clados).
ANAGÊNESE
Chama-se anagênese (do grego: ἀνά = ana = sobre, para cima + Γένεσις = gênese = geração) ao acúmulo de de mudanças (genética, via mutação) que uma população sofre ao longo do tempo, originando uma espécie com características diferentes (evolução gradual). A anagênese é também conhecida como evolução gradual ou mudança filética (segundo alguns pesquisadores). Nessa modalidade de evolução a população ancestral vai acumulando variações genéticas a ponto de, com o tempo ser totalmente distinta da população original, justificando assim a designação de uma nova espécie totalmente distinta. Assim, toda a população atual é diferente da população ancestral e esta esta extinta, sobrevivendo apenas a nova população.
Abaixo vemos uma representação gráfica dos cinco reinos (propostos por Whittaker, 1969) como um caso de cladogênese de taxons superiores.
Abaixo vemos uma representação gráfica dos cinco reinos (propostos por Whittaker, 1969) como um caso de cladogênese de taxons superiores.

SISTEMÁTICA
Cladograma é uma representação gráfica de uma hipótese sobre
o padrão de relações filogenéticas, (i.e., de parentesco ao longo do tempo) de
organismos pertencentes a linhagens diferentes. Através delas, busca-se
reconstruir os eventos de cladogênese, ou seja, da divisão de uma linhagem em
duas que ocorreram durante a evolução.
Nos cladogramas os nós ou nodos (pontos de bifurcação)
representam os ancestrais comuns às duas linhagens. Os cladogramas são feitos a
partir da análise de conjuntos de caracteres (morfológicos ou moleculares),
onde certos caracteres compartilhados (apomórficos) ditos derivados (que se
originaram a partir de modificações nos caracteres anteriores, antigos ou
primitivos (caracteres plesiomórficos) de organismos ancestrais, primitivos.
As sinapomorfias são caracteres homólogos apomórficos
compartilhados por mais de um táxon. É a situação que se observa quando dois ou
mais táxons apresentam o mesmo caractere e este representa uma forma derivada
apomórfica frente à outra ancestral plesiomórfica. Ex.: ovo amniótico, táxons
que apresentam: répteis, aves e mamíferos.
Os diversos cladogramas produzidos pela analise filogenética
através da polarização dos estados dos caracteres em primitivos
(plesiomórficos) e derivados (apomórficos), das diversas características
utilizadas no processo; são então comparados, em geral, utilizando-se a
parcimônia (menor número de eventos de cladogênese, ramificação) como critério de escolha para “o melhor”
cladograma. Podem-se propor árvores de consenso e mais modernamente adotar-se
outros critérios para a construção dos cladogramas, por exemplo baseados em
verossimilhança.
Através destes cladogramas reconstruímos as relações de
parentesco entre os seres vivos, permitindo-nos estudar melhor sua evolução,
além de propor sistemas de classificação naturais muito menos arbitrários e
cientificamente testáveis.
Sua aplicação a microrganismos e vírus também nos permite
estudar a evolução da patogenicidade e virulência destes seres, possibilitando-nos
testar hipóteses e utilizar outros métodos para reconstruir estados ancestrais
destes organismos e até de suas moléculas, o que pode ser valioso para pesquisa
de medicamentos e profilaxia.
Neste texto encontramos varias palavras novas:
Apomorfia
Plesiomorfia
Sinapomorfia
Vamos definir cada um deles para que possamos entender melhor a ciência da classificação.
PLESIOMORFIA
caracteres primitivos, ancestrais
Do grego; Πλησι, Πλησις = plesis: próximo, trazer para perto, associado a, seguir + μορφη = morphe: forma (literalmente forma antiga presente; característica ancestral, primitiva).
É uma característica ancestral, primitiva que vários grupos possuem, deste modo não define nenhum um grupo. É uma característica ancestral (primitiva) herdada de ancestrais distantes no tempo. É um termo empregado em cladística para designar uma característica considerada ancestral (primitiva) que ao longo do tempo foi modificada a outra mais recente (através da mutação, seleção, deriva e fluxo gênico) dentro de uma linhagem.
Para se estabelecer uma plesiomorfia é necessário a comparação da linhagem estudada com um grupo externo (uma linhagem supostamente aparentada filogeneticamente daquela em estudo ou em questão) para se detectar o grupo mais primitivo, mais ancestral. Desta forma, consideramos PLESIOMÓRFICOS os caracteres apresentados pelo grupo primitivo, ancestral.
Ex.:
ausência de vértebras = plesiomorfia
Presença de vértebras = apomorfia

Filogenias mostrando a terminologia usada
para descrever diferentes padrões de estados
de características ancestrais e derivados.
SINAPOMORFIA
caracteres derivados, evoluídos
Do grego: ἀπό: apo = distante, remoto, longe, à distância + μορφή: morphé = forma. Distante da forma antiga, ancestral, primitiva = novidade evolutiva.
Característica nova ou derivada, evoluída, presente apenas em um grupo, e somente num grupo. As apomorfias são deduzidas, estabelecidas a a partir de estudos de anatomia comparada. As características escolhidas devem ser homólogas (i.e., devem ter a mesma origem embrionária). É uma característica presente apenas num táxon, é o que define aquele táxon. A seguir vemos um cladograma mostrando as características apomórficas que definem cada um dos táxons do filo Chordata (cordados).


SINAPOMORFIA
É uma novidade evolutiva compartilhada por todos os descendentes de um ancestral comum (é uma novidade evolutiva para aqueles táxons derivados de um ancestral comum). Do grego σύν: sin = com, junto + ἀπό: apo = a distante, remoto, longe + μορφή: morphé = forma.
São características
homólogas apomórficas compartilhadas por mais de um táxon. É a situação que se
observa quando dois táxons apresentam o mesmo caractere e este representa uma
forma derivada apomórfica frente a outra ancestral plesiomórfica; ex.:
mandíbula: táxons que apresentam mandíbula: peixes, anfíbios, répteis, aves e
mamíferos). Apenas as sinapomorfias constituem argumentos válidos em favor da
monofilia (origem monofilética de grupos) de grupos de táxons que as
compartilham. As sinapomorfias, assim, são o fundamento para a classificação
filogenética (baseado no parentesco) dos seres vivos. Graças às sinapomorfias
se definem os grupos monofiléticos segundo Willi Hennig. A seguir vemos uma árvore filogenética que mostra os caracteres apomórficos e sinapomórficos do táxon Primata.

CONCEITOS IMPORTANTES
ESPÉCIE
Grupo de indivíduos semelhantes
(no nível morfológico e funcional, e bioquímico);
idêntico cariótipo (mesmo número cromossômico)
que vivem numa mesma área geográfica,
capazes de reprodução (cruzam-se) entre si,
originando descendentes viáveis e férteis,
e que estão isoladas reprodutivamente de outros grupos.
POPULAÇÃO
Conjunto de indivíduos da mesma espécie que vivem numa mesma área, ao mesmo tempo e que podem trocar genes entre si.
CLADOGÊNESE
Chama-se cladogênese (do grego: Κλάδος = clados= ramo + Γένεσις = gênese= gerar, origem, nascimento) o conjunto de processos que promovem a especiação, i.e., a separação de uma população em duas ou mais e sua subsequente diferenciação genética e morfológica o que leva ao surgimento de novas espécies (equilíbrio pontuado). Ex. no arquipélago Hawaino, onde colonizadores (pássaros e moscas das frutas) viajaram enormes distâncias via corrrentes marinhas e de vento, chegando aos inúmeros vales de muitas ilhas) se estabelecendo nesses habitats e se diversificando e com o tempo originaram novas espécies (clados).
ANAGÊNESE
Chama-se anagênese (do grego: ἀνά = ana = sobre, para cima + Γένεσις = gênese = geração) ao acúmulo de de mudanças (genética, via mutação) que uma população sofre ao longo do tempo, originando uma espécie com características diferentes (evolução gradual). A anagênese é também conhecida como evolução gradual ou mudança filética (segundo alguns pesquisadores). Nessa modalidade de evolução a população ancestral vai acumulando variações genéticas a ponto de, com o tempo ser totalmente distinta da população original, justificando assim a designação de uma nova espécie totalmente distinta. Assim, toda a população atual é diferente da população ancestral e esta esta extinta, sobrevivendo apenas a nova população.
OU
ANAGÊNESE
É a evolução progressiva de espécies que envolve uma mudança na frequência genética de uma população inteira em oposição a um evento de ramificação cladogenética. Quando um número suficiente de mutações atingem a fixação numa população de tal maneira que existe uma diferença significativa em relação à população ancestral, uma nova espécie pode ser designada com um novo nome. O ponto chave: toda a população é diferente da população ancestral, de tal forma que a população original pode ser considerada extinta. Portanto na ANAGÊNESE, a população vai se modificando gradativamente através de mutações que se fixam (tornam-se estáveis) em função de contínuas alterações nas condições ambientais, o que resulta em uma população tão diferente da original que pode ser considerada uma nova espécie.
CLADOGÊNESE
Processo evolutivo que gera ramificações ou clados (do grego: klados = κλάδος = ramo) ou (diversificação) nas linhagens existentes de organismos ao longo de sua história evolutiva e implica obrigatoriamente em especiação biológica.
Portanto, as novas espécies se formam por irradiação adaptativa, isto é, a partir de grupos que se isolam da população original e se adaptam a diferentes condições ambientais, divergindo ao longo do tempo.

ANAGÊNESE E CLADOGÊNESE
ESPECIAÇÃO
é o processo pelo qual uma espécie divide-se em duas ou mais espécies, que, então, podem evoluir em diferentes linhagens. A etapa crítica na formação de uma nova espécie é a separação do conjunto gênico da espécie ancestral em dois conjuntos separados (fluxo ou deriva).
Posteriormente, em cada conjunto gênico isolado (que seguem histórias evolutivas distintas), as frequências dos genes podem mudar como resultado das ações de forças evolutivas (mutação, seleção, deriva e fluxo). Durante esse período de isolamento, caso diferenças significativas forem acumuladas, as duas populações podem não mais trocar genes. O que forma espécies isoladas, que mesmo em contato com outros conjuntos de indivíduos não há mais fluxo gênico.

CLADOGRAMA
Cladograma (do grego: klados = κλάδος = ramo) é um diagrama usado em cladística e taxonomia que mostra as relações (filogenéticas ou genealógicas) entre táxons terminais, seja em nível de espécie ou grupos supra-específicos, formando grupos monofiléticos evidenciados por sinapomorfias, apomorfias e plesiomorfias, indicando uma história em comum, não necessariamente uma ancestralidade direta. Apesar de terem sido tradicionalmente obtidas principalmente por caracteres morfológicos, as sequências de DNA e RNA e a filogenética computacional são agora normalmente usados para gerar cladogramas. O resultado final de uma análise cladística é apresentado na forma de uma árvore ou cladograma, um dendograma (esquema gráfico) que expressa hipóteses de relações filogenéticas entre táxons.

Cladograma mostrando todos os animais com simetria bilateral.
APOMORFIA
Apomorfia (do grego, απο, "longe de" e μορφη, "forma") é o termo designado para definir uma característica mais recente derivada de uma característica primitiva de uma espécie ancestral.
Uma condição é dita como sendo apomórfica em relação a outra; por exemplo, a presença de carioteca em determinadas células é apomórfica à sua ausência.
SINAPOMORFIA
Sinapomorfia é um termo derivado das palavras gregas: σύν = syn = com, na companhia de, jundo à, junto com; Apo= ἀπό = longe de; e μορφή = morphe = forma.
Sinapomorfia (novidade evolutiva), termo que designa caracteres homólogos apomórficos compartilhados por dois ou mais táxons. É a situação que se observa quando dois táxons apresentam o mesmo caractere, e este representa uma forma derivada apomórfica frente a outra ancestral plesiomórfica.
PLESIOMORFIA
Plesiomorfia (caracteres primitivos) (do grego, πλησιος, "próximo a" e μορφη, "forma") é um termo empregado em cladística para designar uma característica considerada primitiva que foi modificada a outra mais recente dentro de uma linhagem. Para o estabelecimento das plesiomorfias, é necessária a comparação de uma linhagem estudada com um grupo externo (uma outra linhagem supostamente aparentada filogeneticamente daquela em questão) para verificar o grupo supostamente mais primitivo. Daí, consideram-se plesiomórficos os caracteres apresentados por este grupo.


Cladograma imaginário. A máscara amarela é uma plesiomorfia para cada espécie viva mascarada, por ser ancestral. É também uma simplesiomorfia para eles. Mas para as quatro espécies vivas como um todo, é uma apomorfia porque não é ancestral para todas elas. A cauda amarela é uma plesiomorfia e simplesiomorfia para todas as espécies vivas.
Imaginary cladogram.The yellow mask is a plesiomorphy for each living masked species, because it is ancestral. It is also a symplesiomorphy for them. But for the four living species as a whole, it is an apomorphy because it is not ancestral for all of them. The yellow tail is a plesiomorphy and symplesiomorphy for all living species.(W)

Colour pattern diversity of Heliconius numata (top two rows), and H. melpomene
(third row) with its co-mimic H. erato (bottom row).


Padrões de coloração das asas de Heliconius erato phyllis (Fabr. 1775) segundo Ramos (2010)

Heliconius erato phyllis (Fabricius, 1775)
(Fonte: A. Carosia, 2017)
HOMOLOGIA E ANALOGIA
HOMOLOGIA
Homologia (do grego ομοως, "igualmente" e λογος, "ciência", "razão") é o estudo biológico das semelhanças entre estruturas de diferentes organismos que possuem a mesma origem ontogenética e filogenética (evolutivas). A homologia tem sido uma forte evidência em favor da Teoria da Evolução, pois ela sugere ancestralidade comum entre organismos diferentes possuindo estruturas frequentemente semelhantes com a mesma origem embriológica. Por exemplo, as nadadeiras ventrais dos peixes e os membros dos mamíferos.
ANALOGIA
Palavra usada para designar uma comparação (do grego: αναλογία – analogia, "proporção matemática") também conhecido por comparação. Exemplo:
Órgão Análogo é aqueles órgãos que desempenham a mesma função em espécies distintas embora tenham origens embrionárias diferentes, ou seja, as células se diferenciam de folhetos embrionários distintos (mesoderma, endoderma e ectoderma), representando apenas semelhança morfológica entre estruturas, em função de mecanismos adaptativos correlacionados à execução requerida pelo mesmo, por exemplo, as asas das aves e dos insetos, diferentes quanto à origem, mas adaptadas ao vôo.

Órgãos Homólogos, são aqueles que
possuem a mesma origem embrionária e desenvolvimento semelhante em diferentes
espécies, embora em alguns casos possa exercer funções diferentes em diferentes
espécies, como os membros anteriores de vertebrados terrestres: o braço do ser
humano, a pata dianteira de um gato, a nadadeira de uma baleia e as asas de um
morcego.


FILOGENIA
Em biologia, filogenia é o estudo das relações evolutivas entre grupos de organismos (por exemplo, espécies, populações), que é inferido por meio do sequenciamento de dados moleculares e matrizes de dados morfológicos (características morfológicas). O termo filogenética deriva do termos grego Phyle (φυλή) e Filon (φῦλον), que significa: "tribo" e "raça", e o termo genético (γενετικός), que significa "em relação ao nascimento", da gênese (γένεσις) "origem" ou "nascimento". O resultado dos estudos filogenéticos é a história evolutiva dos grupos taxonômicos, ou seja sua filogenia, e é representado por meio de árvores filogenéticas e cladograma.


A pata pentadáctila é um estado ancestral de todos os tetrápodes (que surgiu muito provavelmente de um ancestral que apresentava patas com maior numero de dedos. Ao se adaptar a novos ambientes o numero de dedos foi diminuindo ate se fixar em cinco, constituindo o padrão ancestral de todos os tetrápoda.
Classificação na atualidade
Segundo Lopes e Chow Ho (s/d) há duas escolas principais que se pautam no princípio evolutivo central de descendência com modificação: a evolutiva e a filogenética ou cladística. Ambas partem de um aspecto simples, porém fundamental: organismos com relação de parentesco próxima são mais semelhantes que organismos com relação de parentesco relativamente mais distante. Isto porque parentes próximos tendem a herdar características que estavam presentes em um ancestral em comum.
É fácil notar como irmãos parecem mais entre si do que quando comparados com outros parentes mais distantes ou outras pessoas. Essas escolas divergem, no entanto, no modo como interpretam as relações de parentesco.
Na escola evolutiva, foi partindo desta simples observação que foram desenvolvidas classificações buscando simplesmente agrupar organismos semelhantes que reflitam o maior ou menor grau de parentesco. Para tal, os pesquisadores procuraram observar caracteres que pudessem de alguma forma auxiliar na identificação dos seres mais aparentados, observando peculiaridades ou traços discretos nos organismos como, por exemplo, estruturas, morfologia, citologia, embriologia, etc.
É importante notar dois aspectos principais neste tipo de escola de classificação evolutiva. O primeiro é que as decisões sobre quais caracteres são importantes ou não é feita sem nenhum método objetivo e replicável (repetir o experimento com outros caracteres usando o mesmo protocolo) (aspecto este fundamental em qualquer disciplina científica), e o segundo é que ela não permite a utilização de muitos caracteres simultaneamente.
O fato da escolha da importância dos caracteres ser relativamente arbitrária, dependendo do pesquisador ou dela ser mais evidente e fácil de analisar, tornou-se subsequentemente uma questão central na biologia, pois eles não necessariamente poderiam refletir as proximidades de parentesco. Todos os caracteres têm a mesma importância? Se não tem a mesma importância, quais caracteres devem ser utilizados em sistemas de classificação?
Estas questões só começaram a ser resolvidas com o desenvolvimento, do conceito de
homologia. O curador do Museu de História Natural de Londres, Richard Owen (biólogo, anatomista e palentólogo), foi um
dos grandes responsáveis pelo desenvolvimento desse conceito.

Rochard Owen (1804-1892)
Ele argumentava que existiam
tantas semelhanças entre os membros anteriores de diferentes animais vertebrados como a
nadadeira de uma foca e a mão de um ser humano, que as estruturas deveriam ser derivadas
de uma única estrutura presente no ancestral comum de todos aqueles animais.
Este conceito
entrelaça de maneira elegante e definitiva as observações provenientes da anatomia comparada com o pensamento evolutivo que começava a se desenvolver ao final do século XIX.
Este
grande desenvolvimento intelectual resolve a primeira das questões: quais caracteres devem
ser utilizados ao fazer reconstruções evolutivas? Os caracteres homólogos, por refletirem
ancestralidade comum e, portanto, a herdabilidade de caracteres, são os melhores candidatos
para a reconstrução evolutiva. (Lopes & Chow Ho, s/d).
No entanto, ainda resta um problema de ordem prática.
Como determinar quais caracteres são homólogos? Que método deve ser utilizado para que possamos, de maneira objetiva e replicável, diferenciar os caracteres homólogos?
Por muito tempo, não existiam métodos analíticos para determinar a validade dos caracteres homólogos, e assim, eram consideradas racionalizações discursivas, ou seja, autoridades no assunto estudavam os diversos caracteres a fim de determinar a partir de sua opinião quais deveriam ser utilizados para reconstrução evolutiva e/ou classificação.(Lopes & Chow Ho, s/d)
No entanto, nas décadas de 1950 e 1960, o entomólogo alemão Emil Hans Willi Hennig propôs uma teoria capaz de lidar com todos os caracteres gerados pelos morfologistas de uma vez só, e ainda determinar através de um meio analítico quais seriam homólogos. Sua abordagem, hoje conhecida como cladística ou sistemática filogenética, revolucionou o campo da sistemática. (Lopes & Chow Ho, s/d)
Atualmente, a técnica é utilizada não somente para reconstruções históricas de parentesco entre os organismos, mas também como ferramenta preditiva em estudos epidemiológicos (maiores detalhes sobre isso será tratado ao final do tópico).(Lopes & Chow Ho, s/d).
Na sistemática filogenética, (Mazzarolo, 2005, Lopes & Chow Ho, s/d) entende-se que um caráter, eventualmente, poderá ser modificado na descendência, passando a se apresentar com variações, que serão subsequentemente, herdadas nas próximas gerações.
Desta maneira, o caráter está presente no ancestral exclusivo de todos os herdeiros, e também em todos os herdeiros, mas nestes com uma modificação ou variação.
Essa nova variação ou novo estado do caráter é considerado uma condição derivada, ou seja, surgiu a partir da modificação no estado do caráter previamente presente na linhagem ancestral.
A condição derivada tem o potencial de servir como determinante para definir um novo grupo e é chamada apomorfia (do grego apó = longe de; morphě = forma) no paradigma moderno.
Uma apomorfia pode ser exclusiva de apenas um grupo, sendo chamada nesse caso de autapomorfia (do grego autos = (eu) mesmo, próprio; morphě = forma), ou compartilhada por dois ou mais grupos, chamada sinapomorfia (do grego sýnapsis = ação de juntar; morphě = forma).
Há, no entanto, casos em que o caráter é herdado sem modificação, falando-se em estado plesiomórfico (do grego plesios = vizinho (à forma ancestral), significando próximo; morphé = forma) e não serve para definir um novo grupo.
Quando esse estado plesiomórfico é compartilhado por mais de um agrupamento é chamado simplesiomórfico.
As analogias, ou seja, caracteres semelhantes que surgem em linhagens não aparentadas, devem ser discriminadas nas análises filogenéticas, pois elas podem gerar interpretações errôneas sobre relações de parentesco entre grupos de organismos.
Esses caracteres análogos devem ser identificados para evitar que equivocadamente sejam usados para unir grupos não relacionados. Caracteres análogos são interpretados na filogenética como um tipo de homoplasia (do grego homós = semelhante, igual; plásis = ação de modelar, dar feição): semelhança estrutural decorrente de paralelismo ou convergência evolutiva, e não de ancestralidade comum.
Plesiomorphic is derived from two Greek roots that mean near or close to (plisiazo, πλησιάζω) and form (morfi, μορφή). The reference is to a character that is close to the primitive condition.





CLASSIFICAÇÃO FILOGENÉTICA OU SISTEMÁTICA CLADÍSTICA
Apomorphic is derived from two Greek roots that mean from or off (apo, από) and form (morfi, μορφή). The reference is to a character or state that is removed from the primitive condition. That is, it is a derived condition. An apomorphic character is one that is "advanced" or derivado from a primive or ancestral state.
Consider the example of Urodacus yaschenkoi (an australian scorpion) below. The first walking leg has the more primitive or plesiomorphic state of both claws being almost equal in size. The fourth walking leg, however, has a derived state in which the first claw is greatly enlarged and the second claw is reduced.

O processo evolutivo
A Evolução é entendida como sendo repetições intercaladas de dois sub-processos: a anagênese e a cladogênese.(Mazzarolo, 2005).
La divergence adaptative (différenciation des populations associée à l’adaptation à une nouvelle niche écologique) est un processus clé de la biologie évolutive. L’étude de ce processus est essentielle pour comprendre l’adaptation des organismes à leur environnement et la façon dont la biodiversité se développe. La domestication est un cas de divergence adaptative sous une forte pression humaine et à l’échelle temporelle humaine. Charles Darwin a utilisé des pigeons pour illustrer le processus et le pouvoir de sélection dans des formes de rendement radicalement différentes de la forme sauvage ancestrale en comparant les pigeons sauvages et domestiqués. (Ropars et alii, 2020).
A divergência adaptativa (diferenciação de populações associada à adaptação a um novo nicho ecológico) é um processo chave na biologia evolutiva. Estudar esse processo é essencial para entender como os organismos se adaptam ao meio ambiente e como a biodiversidade se desenvolve. A domesticação é um caso de divergência adaptativa sob forte pressão humana e em uma escala temporal humana. Charles Darwin usou pombos para ilustrar o processo e o poder de seleção em formas de produção dramaticamente diferentes da forma selvagem ancestral, comparando pombos selvagens e domesticados.
Desde então, a domesticação tem sido usada como um bom modelo para o entendimento da adaptação, pois envolve uma seleção forte e recente de características que podem ser facilmente identificadas. Essa seleção forte e recente por humanos geralmente deixa traços de adaptação nos genomas que são mais fáceis de detectar do que aqueles deixados pela seleção natural. A comparação de populações domesticadas e não domesticadas nos níveis fenotípico e genômico pode identificar a arquitetura genética de características adaptativas e mecanismos genômicos permitindo rápida adaptação e diferenciação, como foi demonstrado para milho e cães, por exemplo.(Ropars et alii, 2020).
A anagênese é a parte do processo evolutivo onde a forma se modifica ao longo do tempo. Ou seja, é na anagênese que as características de uma determinada espécie podem se modificar atraveés da seleção natural, evoluir, com o decorrer do tempo.
Por exemplo, durante a anagênese de uma espécie A, a cor negra de seus olhos pode ser substituída pela cor vermelha ao longo de algumas gerações. Esse processo se daria da seguinte forma teórica simplificada: imaginemos que num determinado tempo inicial T1 todos os indivíduos da espécie A apresentem os olhos com a cor negra. (Mazzarolo, 2005)
Em um tempo seguinte T2, através de uma mutação, aparecem alguns indivíduos com os olhos com a cor vermelha. Imaginemos, então, que esse novo estado do caráter cor do olho seja adaptativo, i.e., confira certa vantagem competitiva (por alimento, por parceiros reprodutivos, etc.) aos indivíduos que apresentam os olhos vermelhos. Nesse caso, este novo estado (ou estado derivado) será selecionado pela pressão ambiental, e gradativamente irá aumentando o número de indivíduos com olhos vermelhos até ocorrer o desaparecimento dos indivíduos de olhos negros nessa espécie.

Em T1 (início da observação) uma população natural apresenta variações. Em T1 uma surge uma mutação. Essa mutação produz um novo alelo na população. Indivíduos que apresentam esse alelo novo possuem vantagem seletiva sobre os demais alelos. O olho vermelho é mais adaptado nesse ambiente. Em T3, podemos observar que essa variante perfaz toda a população.
Então, ao final da anagênese da espécie A, no tempo T3, todos os seus indivíduos apresentarão os olhos vermelho.
É importante notar que, apesar da modificação da forma ao longo do tempo, SEMPRE existiu uma única espécie, a espécie A, ou seja, não ocorreu aumento da diversidade, i.e., não houve especiação. (Mazzarolo, 2005)
Para que ocorra o aumento da diversidade é necessário que essa espécie A (agora chamada espécie ancestral) origine duas espécies descendentes, B e C, a partir de um evento de cladogênese. Ou seja, a cladogênese é o evento de "quebra" de uma espécie ancestral em duas espécies descendentes (especiação), gerando um aumento na diversidade (numa área onde antes havia apenas u ma espécie agora há duas espécies).
Essa quebra deve interromper o fluxo gênico (crossing over e segregação independente) entre os indivíduos das espécies descendentes, agora isoladas uma da outra. (modif. de Mazzarolo, 2005)
Ou seja, no nosso exemplo acima, não deve mais ocorrer fluxo gênico entre os indivíduos da espécie descendente B com os indivíduos da espécie descendente C e vice-versa.
As espécies descendentes B e C estão agora isoladas entre si e são consideradas duas espécies distintas (i.e., boas espécies). Provavelmente, na maioria dos casos, os eventos de cladogênese são provocados por um evento espacial ou geográfico, ou seja, a partir fragmentação da área geográfica inicial, ocorre a divisão da espécie (ou das espécies) que vive nela, originando as espécies descendentes. (Mazzarolo, 2005)
A fragmentação geográfica que gerou um aumento da diversidade (especiação) mais conhecida é a fragmentação da Pangea nos diversos continentes atuais; outros exemplos são: surgimento de um rio que separa uma população em duas, orogenia ou o surgimento de montanhas onde antes havia uma unica população ou espécie, surgimento de vales por ex., nos Andes ou nos Alpes europeus, surgimentos de ilhas ou arquipélagos (Galápagos e Hawai entre outros), transgressões marinhas gerando ilhas, onde antes existiam apenas montanhas. (modif. de Mazzarolo, 2005).

Sinapomorfias e plesiomorfias na evolução dos mamíferos.
(Modificado de Dictionary)
A evolução das características
Uma vez que compreendemos o processo gerador da biodiversidade (especiação), devemos agora procurar compreender o método proposto pela Sistemática Filogenética ou Cladistica para a recuperação da informação filogenética (ao menos dos padrões obtidos pela observação na natureza).
Como recuperar a história evolutiva de um grupo taxonômico (táxom) através do estudo de suas características atuais?
Inicialmente devemos abordar o conceito de homologia.

Homologia (Lovo s/d)

Homologia
(Lovo s/d)
São considerados caracteres homólogos aqueles que tem a mesma origem ontogenética (embrionária), podendo ou não apresentar a mesma função. Apresentar homologia significa possuir genes que produzem esses órgãos ou características.
Por exemplo, são considerados homólogos o braço direito de um homem e o de um chimpanzé, a asa de uma gaivota, a asa de um morcego, bem como o braço direito de um anfíbio e a perna anterior direita de um cavalo.
Ao compararmos estruturas homólogas estamos analisando estruturas que tiveram origens na mesma estrutura ontogenética, ou seja, são a "mesma" estrutura. (modif. de Mazzarolo, 2005).
Não poderíamos, por exemplo, comparar o braço direito de um homem com a perna posterior direita de um cavalo, pois esta perna do cavalo não é homóloga ao braço direito e sim à sua perna direita do homem (membros posteriores).
Aparentemente, encontrar homologia entre diferentes estruturas parece mais fácil do que realmente é, e um engano nessa fase inicial do estudo comprometerá toda a análise sobre a evolução da estrutura e, conseqüentemente, toda a inferência filogenética decorrente desse processo.
Após construirmos uma hipótese confiável sobre a homologia das estruturas a serem comparadas, passamos para a etapa seguinte. (Mazzarolo, 2005)
Nesse momento, a partir dos dois estados diferentes encontrados para a mesma estrutura, devemos buscar compreender como foi a série de transformação desse caráter, dessa estrutura, ou seja, devemos propor uma hipótese a respeito de como essa estrutura, evoluiu, de como ela se modificou ao longo do tempo. (Mazzarolo, 2005)
Devemos nos lembrar que as modificações nas estruturas ocorrem durante o processo de anagênese. Desse modo, lembrando dos exemplos anteriores, devemos conseguir recuperar a informação de que os olhos negros se transformaram em olhos brancos, ou devemos conseguir inferir se a perna anterior do cavalo se transformou no braço do homem ou se o que ocorreu foi o contrário, a partir de um braço ocorreu a transformação em perna anterior.
Como vimos, os estados dos caracteres podem ser diferenciados em ancestrais, mais antigos, denominados plesiomórficos, ou plesiomorfias, e derivados, mais recentes, denominados apomórficos, ou apomorfias. Uma apomorfia deriva, modifica-se, a partir de uma plesiomorfia.(Mazzarolo, 2005)
Agora temos duas novas perguntas a serem respondidas:
1. Para que distinguir entre uma apomorfia e uma plesiomorfia?
2. Como distinguir entre uma apomorfia e uma plesiomorfia?
Para respondermos a primeira devemos relembrar como é compreendido o processo evolutivo. Ele é um conjunto de anagêneses intercaladas por um conjunto de cladogêneses.
Se as duas espécies descendentes recebem o estado modificado do caráter (apomorfia) de sua espécie ancestral comum e exclusiva, o pensamento inverso também é válido.
Se duas espécies apresentam estado derivado do caráter (apomorfia), isso sugere que elas o receberam de uma espécie ancestral comum e exclusiva, onde o estado primitivo se modificou, durante sua anagênese, no estado derivado.
Este estado derivado compartilhado por duas (ou mais) espécies é denominado sinapomorfia.(Mazzarolo, 2005)
Então, a identificação de sinapomorfias sugere a existência de ancestrais comuns exclusivos, o que indica maior relacionamento de parentesco entre os táxons que a compartilham. Ou seja, podemos reconstruir o padrão filogenético de um grupo a partir da identificação de estados de caráter derivados compartilhados (sinapomorfias).
Para respondermos a segunda pergunta que lida coma distinção entre uma apomorfia e uma plesiomorfia, devemos estudar a embriologia e dos táxons a serem comparados e sua anatomia, embriologia, fisiologia comparados. Assim, podemos distinguir dois métodos que podem ser utilizados para o que chamados de Polarização de caráter.
1) Método ontogenético
2) Método do grupo externo ou out group.
O estado desse caráter no grupo externo será inferido como o estado ancestral ou plesiomorfico e o estado observado no grupo em estudo será o estado derivado ou apomorfico.

CLASSIFICAÇÃO FILOGENÉTICA OU SISTEMÁTICA CLADÍSTICA
Em 1859, Charles Darwin publicou a obra que inaugurou o pensamento evolutivo moderno (cf. Mayr & Bock, 2002; Gould, 2002; Bowler, 2003; Ruse & Travis, 2009). No ano anterior, já haviam sido divulgados os artigos que tratavam do que viria a tornar-se a teoria central das ciências biológicas, ou seja, a evolução via seleção natural (cf. Darwin, 1858; Wallace, 1855, 1858). Para Mayr (1998; 2005), o trabalho de Darwin (1859) compreende cinco teorias independentes, mas relacionadas:
(1) a evolução propriamente dita,
(2) a descendência com modificação,
(3) o gradualismo,
(4) a especiação (multiplicação das espécies) e
(5) a seleção natural.
A percepção de um mundo orgânico em constante mudança e a ideia de que todos os organismos compartilham um ancestral comum em algum momento de sua história evolutiva revolucionou o pensamento biológico no século XIX. O objetivo central de Darwin era negar o fixismo das espécies e sua criação em separado (cf. Ruse & Travis, 2009). (Santos e Klassa, 2012)
A introdução da ideia de uma árvore da vida nas ciências biológicas, somada às proposições acerca dos mecanismos pelos quais a evolução ocorria, implicou em reconsiderações fundamentais para as classificações biológicas. A visão ainda vigente de uma scala naturae (escala da natureza) foi paulatinamente substituída pela noção de uma grande e única árvore da vida, uma vez que todas as formas de vida existentes assemelham-se em algum grau (ascendência comum). Coube à sistemática a busca pelos "parentes" mais próximos a todos os grupos e a reconstrução dos seus ancestrais comuns (cf. Mayr, 1998).
Segundo Darwin, "o arranjo dos táxons deve ser estritamente genealógico, mas a quantidade de diferenças em vários ramos ou grupos, ainda que unidos no mesmo grau em sangue de seu progenitor ancestral, pode diferir enormemente e isso deve ser considerado no escalonamento. Os graus de modificação que os diferentes grupos sofrem devem ser expressos em diferentes categorias" (Darwin, 1859, p. 479-80).
A despeito do trabalho de Darwin (1859), Wallace (1855) tem prioridade em explicar evolutivamente a existência das categorias taxonômicas supragenéricas. Ele foi o pioneiro em temporalizar as classificações, explicando a quantidade de diferenças morfológicas por diferentes origens no tempo. Os dois critérios citados por Darwin, a genealogia e o grau de semelhança, foram incorporados pelas classificações biológicas como peças-chave de suas análises, dado o interesse pelo entendimento da história evolutiva dos organismos. (Santos e Klassa, 2012).
Inúmeros sistematas voltaram-se para o estudo das relações de parentesco entre os organismos, e a escola conhecida como sistemática ou taxonomia evolutiva tornou-se a mais popular nas décadas de 1930-1950 (cf. Mayr et al., 1953; Mayr & Provine, 1980). Para Mayr (2008), ela trazia a única representação passível de satisfazer ambos os critérios estabelecidos por Darwin para uma classificação consistente.(Santos e Klassa, 2012).
Essas classificações reuniam os grupos e os representavam em um diagrama de árvore, apresentando as relações de parentesco entre as linhagens e suas taxas de divergência morfológica (demonstradas respectivamente pela proximidade dos ramos e diferença de tamanhos dos mesmos).
O objetivo era seguir à risca a premissa de Darwin ao determinar que o conhecimento da filogenia dos grupos por si só era insuficiente na proposição de uma classificação, pois, ainda que unidos por caracteres homólogos, os ramos evoluíam e divergiam em taxas muito diferentes (Mayr et al., 1953; Mayr, 2008). Tal desigualdade deveria ser levada em conta na determinação do posicionamento hierárquico. Em outras palavras, para que um táxon fosse considerado um grupo natural monofilético, deveriam ser analisadas também as características adaptativas conquistadas ao longo da história evolutiva do grupo, atribuindo-lhes valores diferenciados. Assim, os atributos relacionados às grandes adaptações possuiriam um valor evolutivo/adaptativo superior às características compartilhadas que uniriam os ramos, evidenciando uma disparidade morfológica, fisiológica ou comportamental, tal que justificaria a elevação do táxon a uma nova categoria hierárquica.(Santos e Klassa, 2012)
Esse método foi amplamente empregado até a metade do século XX, mas, à medida que novos estudos e propostas de classificação foram sugeridos, suas deficiências tornaram-se aparentes.
A incorporação de conhecimentos prévios dos organismos e a pesagem a posteriori de atributos mediante observações a respeito de sua história evolutiva resultavam em hipóteses filogenéticas não consensuais e em um subjetivismo que impedia sua "testabilidade", além de basear-se excessivamente na autoridade do pesquisador (cf. Santos, 2008). A despeito de trabalharem com o mesmo grupo de organismos, raramente dois taxonomistas clássicos assumiam as mesmas condições sobre os caminhos da evolução (cf. Lipscomb, 1998), o que levava à obtenção de hipóteses distintas. (Santos e Klassa, 2012)
Na década de 1950, dois métodos desvincularam os critérios de genealogia e de grau de similaridade, referidos na taxonomia evolutiva, para resolver a falta de rigor dos estudos filogenéticos até então realizados (cf. Hull, 1988; Santos, 2008). A taxonomia numérica, posteriormente chamada apenas fenética, propunha o agrupamento de organismos fenotipicamente semelhantes, isentando o sistemata de conhecimento evolutivo prévio e excluindo de suas atribuições qualquer diferenciação entre os caracteres observados (tais proposições sobre o processo evolutivo acrescentavam ambiguidade às análises e deveriam ser descartadas). (Santos e Klassa, 2012)
Segundo a concepção dos feneticistas, o objetivo das atividades classificatórias seria apenas o de catalogar e facilitar o acesso às informações sobre a diversidade biológica, pois "as estimativas da 'verdadeira' afinidade, no sentido filogenético, são lógica e historicamente posteriores às estimativas de similaridade global" (Cain & Harrison, 1958, p. 86); as classificações deveriam ser elaboradas por métodos estatísticos claros e precisos. Contudo, como apontado por Santos (cf. 2008), a objetividade do método era ilusória, dado que a própria escolha dos caracteres a serem comparados dependia do observador. Desse modo, a fenética desviou-se muito da classificação biológica de fato (cf. Hull, 1988), uma vez que os feneticistas não levavam a evolução em conta para a construção dos seus fenogramas, contrariando, ou ao menos desconsiderando, a ideia darwiniana de que o processo evolutivo é o responsável pelo padrão hierárquico presente de similaridades no mundo natural (cf. Darwin, 1859).
Contrariamente, a sistemática delineada em Hennig (1950) e modificada em Hennig (1966) tinha como base para a definição dos seus agrupamentos naturais a história evolutiva dos grupos.
Se um dos principais preceitos da teoria da evolução dizia respeito à ancestralidade comum, e somente com a reconstrução desses ancestrais é que se poderia inferir com propriedade a história evolutiva dos organismos, fósseis e viventes, era sensato que as classificações biológicas refletissem, antes de qualquer outro aspecto, as relações de parentesco entre os seres vivos e utilizassem o conceito de ancestralidade comum como fundamento para a identificação do monofiletismo. Esse era o mote do método de Hennig.


A metodologia filogenética consistia na observação de certos atributos nos diferentes organismos seguida da formulação de hipóteses de homologia, que seriam então confrontadas posteriormente no chamado teste de congruência (cf. Nelson & Platnick, 1981; Farris, 1983), para o estabelecimento das relações de parentesco. Eram esses caracteres homólogos que sugeririam a ascendência comum. O refinamento da sistemática filogenética em relação à taxonomia evolutiva estava em discriminar caracteres primitivos (plesiomórficos) de derivados (apomórficos), e estabelecer relações de parentesco apenas a partir do compartilhamento dessas apomorfias, a fim de distinguir as homologias das convergências e determinar os grupos monofiléticos (no sentido cladístico), que são os únicos que realmente respeitam o conceito da ancestralidade comum (cf. Santos, 2008). Assim, o sistemata hennigiano identifica um clado natural como a reunião da espécie tronco (ancestral) mais recente e de todos os seus descendentes, determinada a espécie pela presença de caracteres compartilhados exclusivamente pelos descendentes (sinapomorfias), independente do grau de divergência que possa existir. (Santos e Klassa, 2012)
O escalonamento (ou hierarquia) originado por esse método acontece naturalmente, refletindo a história evolutiva e estabelecendo as relações de grupo-irmão entre os grupos-naturais, isto é, os clados. A forma precisa pela qual a análise cladística definiu a relação biológica de parentesco (o táxon A está mais próximo de B em relação a C), ordenando as sinapomorfias de modo a obter uma classificação dos táxons consistente com sua genealogia, calcada no princípio da parcimônia (cf. Hennig, 1966; Farris, 1983), reflete a grande contribuição de Hennig para a sistemática biológica: a formulação de um método propriamente dito, testável, não arbitrário e reflexivo da realidade natural, representada em dendogramas ramificados, chamados cladogramas (que correspondem a conjecturas sobre as relações de parentesco resultantes do processo evolutivo). Em princípio, quaisquer características observadas em organismos possuem o potencial de fornecer evidências de relação de parentesco. No entanto, as evidências mais objetivas são derivadas daqueles atributos que são hereditários e intrínsecos dos organismos, porque eles refletem a continuidade biológica entre ancestral e descendente (cf. Hennig, 1966). A reconstrução dessas relações de parentesco pode ser repetida e testada por conta do sistema congruente e sólido elaborado por Hennig.(Santos e Klassa, 2012)
Emil Hans Willi Hennig (20/abril/1913 – 05/novembro/1976) was a German biologist who is considered the founder of phylogenetic systematics, also known as cladistics. In 1945 as a prisoner of war, Hennig began work on his theory of cladistics, which he published in German in 1950, with a substantially revised English translation published in 1966. With his works on evolution and systematics he revolutionised the view of the natural order of beings. As a taxonomist, he specialised in dipterans (true flies).
Hennig coined the key terms synapomorphy, symplesiomorphy and paraphyly.
He also asserted, in his "auxiliary principle", that "the presence of apomorphous characters in different species 'is always reason for suspecting kinship [i.e., that species belong to a monophyletic group], and that their origin by convergence should not be presumed a priori' (Hennig, 1953). This was based on the conviction that 'phylogenetic systematics would lose all ground on which it stands' if the presence of apomorphous characters in different species were considered first of all as convergences (or parallelisms), with proof to the contrary required in each case." This has been viewed as an application of the parsimony principle to the interpretation of characters, an important component of phylogenetic inference.
He is also remembered for Hennig's progression rule in cladistics, which argues controversially that the most primitive species are found in the earliest, central part of a group's area.

Emil Hans Willi Hennig
(20/abril/1913 – 05/novembro/1976)
(Fonte: Gerd Hennig)
A Cladística (do grego: κλάδος = ramo), também conhecida como sistemática filogenética, é uma escala da Sistemática Biológica baseada no princípio filogenético e que agrupa espécies ou táxons em agrupamentos naturais (gerando classificações hierárquicas ou não) de acordo, unicamente, com hipóteses de relações evolutivas. Segundo os fundamentos da cladística, formulados em grande parte pelo entomólogo alemão Willi Hennig, uma classificação deve sempre expressar as relações evolutiva das espécies, não importando se as espécies são semelhantes ou diferem drasticamente entre si em sua morfologia. Segundo este modelo, em uma classificação cada grupo deve ser obrigatoriamente MONOFILÉTICO, ou seja, possuir todos os descendentes do ancestral comum mais recente do grupo estudado e nada mais. De modo que grupos PARAFILÉTICOS e POLIFILÉTICOS não são permitidos na classificação.
O método CLADÍSTICO é explícito e diretamente conectado à ideia de que o mundo natural ordena-se de forma hierárquica, resultante do processo evolutivo. Não obstante, as classificações filogenéticas são sistemas que permitem não só o armazenamento da informação biológica, levantada para a sua construção, como também a recuperação dessa informação, os grupos monofiléticos representam padrões hierárquicos que carregam consigo a informação sobre a evolução das características das linhagens biológicas consideradas, o que não ocorre nos grupos parafiléticos e polifiléticos, dos quais pode ser depreendida apenas parte da informação usada na construção da classificação.
O monofiletismo é fundamental para a sistemática filogenética e é ele que garante o sucesso do método no que tange à identificação da afinidade natural (evolutiva) entre grupos biológicos. Foi devido ao método de análise cladística das relações de parentesco baseada em padrões evolutivos que os métodos mais antigos e laboriosos, baseados apenas em reuniões de semelhanças em vez de conhecimentos sobre os processos evolutivos, puderam ser repensados.
A cladística forneceu os meios para integrar a enorme quantidade de conhecimento descritivo sobre os seres vivos, gerando uma visão unificada sobre a biodiversidade. Zoologia e Botânica, sob o enfoque filogenético, ganharam um grande dinamismo e nos mostraram como se deu a evolução das características ao longo do tempo.

Arvores Filogenéticas
Pode ser considerada uma arvore genealógica;
Pode englobar diversos níveis de agrupamentos dos seres vivos.
Uma informação que é importante para os pesquisadores é saber quais grupos de seres vivos compartilham um único ancestral comum e exclusivo. n Essa informação possibilita definir um clado válido a ser utilizado na filogenética. n Clado: é um agrupamento que inclui um ancestral comum e todos os descendentes, vivos e extintos desse ancestral (Cladograma).


Homologia: similaridade com origem comum
Método Cladísticos ou baseados em um modelo evolutivo
Calculam árvores para cada caráter e então indicam a melhor arvore através da determinação daquela que requer menor número de mudanças (método da parcimônia) ou aquela mais provável estatisticamente (máxima verossimilhança).
A árvore que melhor explica os relacionamentos entre as seqüências de atributos é denominada cladograma.
Videos recomendados
Diz-se que um grupo de organismos tem origem comum se todos partilham um ancestral. A teoria da origem comum universal postula que todos os organismos em nosso planeta descendem de um ancestral comum, ou de um um pool gênico ancestral.
CARACTERÍSTICAS DERIVADAS
É um estado mais avançado de uma característica anatômica, morfológica etc., presente em um táxon.
ORIGEM COMUM
Segundo Darwin e a Sistemática filogenética de Hennig, todos os seres vivos descendem de um ancestral comum que vivou há muito tempo em nosso planeta, por volta de 3,8 bilhões de anos. Os pesquisadores o chamam de LUCA: Last Universal Common Ancestor.
TÁXONS TERMINAIS
Todos os táxons originados ao longo do tempo de um ancestral comum, são tidos como monofiléticos por definição.
ADENDO
ORIGEM DO CONCEITO DE ESPÉCIE
Como se formam novas espécies? Como se originam novas espécies ?
O fenômeno do surgimento de espécies novas (ou boas espécies) é chamado de especiação. Para que possamos entender como uma espécie nova surge temos que entender antes o que é uma espécie.
O QUE É UMA ESPÉCIE?
A
definição de espécie se alterou ao longo do tempo como resultado da
contribuição de inúmeras outras ciências tais como: História natural, Geologia,
Biologia, Genética, Cladística,
Taxonomia, Sistemática. Desde Platão, Aristóteles, Teofrasto e Santo Agostinho, já vistos, muitos outros propuseram conceitos para
Platão,
Aristóteles, Teofrasto de Eresos, Agostinho de Tagaste,
John Wray, Carl von Linné, Edward Poulton, Theodosius Dobzhanski e Ernst Mayr.
No
século XVII, John Ray (Wray) (1627-1705), filho de um ferreiro, aos 16 anos foi enviado
por seu pai para Cambridge (Cambridge University), onde estudou Grego, Matemática e Humanidades. Hoje é considerado o pai da História natural inglesa. Esse pesquisador foi o primeiro
a elaborar um conceito para espécie.
Usando o critério da fecundidade
(fertilidade), considerava que pertenceriam a mesma espécie, organismos que, através
de cópula, originassem descendentes semelhantes. Suas ideias estão nos
seguintes livros:
Methodus plantarum nova (1682),
Historia generalis plantarum (3 vols. 1686, 1688,
1704),
Catalogus plantarum Angliae
(1670), entre outros inúmeros livros publicados.
No século XVIII, Carl von Linné, (Carolus
Linnaeus), botânico, zoólogo e medico Sueco; através de seus estudos
comparativos propôs a Nomenclatura binomial (classificação científica), sendo
considerado por isso o pai da Taxonomia moderna. Usando um critério de semelhança
morfológica considerou como pertencentes
a uma mesma espécie organismos que apresentassem características morfológicas
idênticas. Esse conceito embora fixista apresenta uma primeira padronização, pois considera a espécie como unidade de classificação. Esse
conceito está exposto em três de seus mais importantes trabalhos: Systema Naturae (1735); Genera Plantarum (1735-1737) e Especies Plantarum (1753).
Embora
de grande valor prático e historicamente importante, este critério foi
rapidamente questionado pois sabe-se que existem organismos muito semelhantes
que pertencem a espécies diferentes (espécies crípticas), e organismos
diferentes que pertencem a uma mesma espécie .
Outros
critérios foram propostos, por ex.: critério genético:
espécie é um grupo de seres vivos geneticamente semelhantes (apresentam o mesmo
pool gênico), o critério ecológico:
uma espécie é um grupo de organismos que apresentam o mesmo nicho ecológico; entre
outros critérios.
Conceito biológico de Espécie:
Edward Poulton, Theodosius Dobzhansky e Ernst Mayr
Um
dos primeiros conceitos biológico de espécie foi desenvolvido por Edward
Bagnall Poulton. Para Poulton o verdadeiro critério de espécie foi a singamia (chamamos singamia ao processo
em que duas células, por exemplo, um espermatozóide e um ovócito se juntam durante
a fertilização), ou compatibilidade reprodutiva, incluindo a compatibilidade
pré e pós-acasalamento. Poulton sugeriu que o cruzamento (singamia) era a
verdadeira natureza das espécies.
Em 1904, Poulton publicou o trabalho intitulado: ”O que é uma espécie?” Este foi a primeira publicação a elaborar o que ficou conhecido como o conceito biológico de espécie, a partir do seu conhecimento sobre mimetismo polimórfico em borboletas Papilio sp (Papilonideos), onde formas masculinas e femininas eram membros da mesma espécie, apesar de serem bastante distintas morfologicamente, pois pertenciam a um mesmo conjunto reprodutivo (população, espécie).
Em 1904, Poulton publicou o trabalho intitulado: ”O que é uma espécie?” Este foi a primeira publicação a elaborar o que ficou conhecido como o conceito biológico de espécie, a partir do seu conhecimento sobre mimetismo polimórfico em borboletas Papilio sp (Papilonideos), onde formas masculinas e femininas eram membros da mesma espécie, apesar de serem bastante distintas morfologicamente, pois pertenciam a um mesmo conjunto reprodutivo (população, espécie).
Outro
pesquisador Theodosius Dobzhansky, publicou um artigo, em 1935, que discutiu o conceito de Poulton. Dobzhansky propôs que uma espécie é um grupo de indivíduos
completamente férteis entre si, mas impedido de cruzamento com outros grupos
semelhantes por suas propriedades fisiológicas, produzindo ou incompatibilidade de pais ou esterilidade do híbrido, ou ambos. Foi
nessa publicação que Dobzhansky desenvolveu o conceito de isolamento pré-zigótico
e o pós-zigótico. Ele incluiu isolamento geográfico como um mecanismo de
isolamento gerador de novas espécies mas
argumentou que apenas os mecanismos de isolamento fisiológico foram importantes
na definição de espécies. Ele chegou a essa conclusão com base em seus trabalhos
com espécies crípticas de mosca das frutas Drosophila
spp.
Ernst
Mayr, biólogo germano-americano, renomado ornitologista, taxonomista e explorador,
apresentou em 1940 e 1942 trabalhos que contribuíram
para a revolução conceitual que culminou na “moderna síntese evolucionária”
(que foi a síntese que ocorreu entre a Genética Mendeliana, a Sistemática e a
Evolução Darwiniana via seleção natural), produzindo-se o que se chama de "Teoria Sintética da Evolução"; através dessa síntese surge um novo
conceito biológico de espécie.
Definido
por Ernst Mayr em 1942, o conceito biológico de espécie está baseado no
isolamento reprodutivo.
“Espécies
são grupos de populações naturais verdadeiramente ou potencialmente intercruzantes,
mas que são isoladas reprodutivamente de outros grupos com as mesmas
características” (Mayr, 1942).
Assim, uma espécie seria definida como sendo o conjunto de indivíduos semelhantes morfologicamente, que vivem em uma dada área que são capazes de real ou potencialmente copularem e produzirem descendestes férteis, e que estão isolados reprodutivamente de outros grupos.
Assim, uma espécie seria definida como sendo o conjunto de indivíduos semelhantes morfologicamente, que vivem em uma dada área que são capazes de real ou potencialmente copularem e produzirem descendestes férteis, e que estão isolados reprodutivamente de outros grupos.
Consequentemente, podemos olhar a espécie como sendo uma evolução das populações a seus ambientes e seu estudo está muito influenciado pela genética de populações. Assim, a especiação (origem de novas espécies) é uma causa do isolamento das populações mendelianas que ao serem impedidas de trocarem genes entre si, se diferenciam e seguem caminhos evolucionários distintos. Por tratar-se de um conceito horizontal (espacial), apresenta limitações: não pode ser aplicado em organismos fósseis, por não se reproduzirem, não pode ser aplicado a organismos com reprodução assexuada uma vez que estes não se produzem via cruzamento (todos são clones, bactérias, fungos e algumas plantas e animais).
Podemos
concluir que não existe um conceito universal (para todos os casos); devendo-se
aplicar os conceitos que melhor expliquem o caso em estudo. Todavia, a espécie
deve ser considerada a unidade de classificação, e a única categoria taxonômica
natural existente, do ponto de vista reprodutivo, genético e ecológico.
VIDEOS RECOMENDADOS
SISTEMÁTICA FILOGENÉTICA
Bibliografia
Bates, H. W. 1862. Contributions to an insect fauna of the Amazon Valley. Lepidoptera: Heliconidae. Trans. Linn. Soc. Lond. 23:495-566.
Beltrán M, Jiggins CD, Brower AVZ, Bermingham E, Mallet M. 2007. Do pollen feeding, pupal-mating and larval gregariousness have a single origin in Heliconius butterflies? Inferences from multilocus DNA sequence data. Biological Journal of the Linnean Society in press.
Beltrán M, Jiggins CD, Bull V, Linares M, Mallet J, McMillan WO, and Bermingham E. 2002. Phylogenetic discordance at the species boundary: comparative gene genealogies among rapidly radiating Heliconius butterflies. Mol. Biol. Evol. 19: 2176-2190.
Benson WW, Brown KS, Gilbert LE 1976. Coevolution of plants and herbivores: passion flower butterflies. Evolution 29, 659-680.
Brower, A. V. Z. 1997 The evolution of ecologically important characters in Heliconius butterflies (Lepidoptera: Nymphalidae): a cladistic review. Zool. J. Linn. Soc. 119, 457-472.
Brower AVZ, and Egan MG. 1997. Cladistics of Heliconius butterflies and relatives (Nymphalidae: Heliconiiti): the phylogenetic position of Eueides based on sequences from mtDNA and a nuclear gene. Proc. R. Soc. Lond. B 264: 969-977.
Brower, L. P., Brower, J. v. Z. & Collins, C. T. 1963. Experimental studies of mimicry 7: relative palatability and Mullerian mimicry among neotropical butterflies of the subfamily Heliconiinae. Zoologica 48, 65-84.
Brown KS, Jr. 1981. The biology of Heliconius and related genera. Ann. Rev. Entomol. 26: 427-456.
Deinert EI. Longino JT, Gilbert LE. 1994. Mate competition in butterflies. Nature 370: 23-24.
DeVries P. J. 1987. The Butterflies of Costa Rica and Their Natural History, Volume I: Papilionidae, Pieridae, Nymphalidae Princeton University Press, Baskerville, USA.
Ehrlich PR, Gilbert LE. 1973. Population structure and dynamics of the tropical butterfly Heliconius ethilla. Biotropica 5: 69-82.
Emsley MG. 1965. Speciation in Heliconius (Lep., Nymphalidae): morphology and geographic distribution. Zoologica NY 50: 191-254.
Gilbert LE. 1972. Pollen Feeding and Reproductive Biology of Heliconius Butterflies. Proceedings of the National Academy of Sciences of the United States of America 69: 1403-1407.
Joron M, Papa R, Beltrán M, Chamberlain N, Mavárez J, Baxter, S, Abanto, M, Bermingham, E, Humphray, SJ, Rogers, J, Beasley, H, Barlow, K, ffrench-Constant, RH, Mallet, J, McMillan, WO, Jiggins, CD. 2006. A conserved supergene locus controls colour pattern diversity in Heliconius butterflies. PLoS Biology Vol. 4, No. 10, e303 doi:10.1371/journal.pbio.0040303
Linnaeus, C. 1758 Systema Naturae, (10th edn., facsimile reprint, 1956). London: British Museum (Natural History).
Mallet J. Singer MC. 1987. Handling effects in Heliconius: where do all the butterflies go? J. Animal Ecology. 56: 377-386.
"Muses." Encyclopedia Mythica from Encyclopedia Mythica Online. http://www.pantheon.org/articles/m/muses.html [Accessed May 22, 2008].
Penz C.M. 1999. Higher level phylogeny for the passion-vine butterflies (Nymphalidae, Heliconiinae) based on early stage and adult morphology. Zoological Journal of the Linnean Society 127: 277-344.
Turner, J.R. 1976. Adaptive radiation and convergence in subdivisions of the butterfly genus Heliconius (Lepidoptera: Nymphalidae). Zoological Journal of the Linnean Society 58(4): 297-308.
www.heliconius.org. Everything about Heliconius.
Chris Jiggins homepage. Genetic linkage mapping of the Heliconius genome to search for regions controlling traits of interest
Los Heliconiini (Lepidoptera, Nymphalidae) de Venezuela . An excellent resource for Venezuelan species.
James Mallet homepage. Information on mimicry and Heliconius in general.
Mathieu Joron homepage.Information about Heliconius numata
Owen McMillan Lab. Mapping colour pattern genes in Heliconius erato.
Robert D. Reed. Evo-devo of Heliconius wing patterns
Insect Flight Research. Butterfly flight , including mimicry in flight patterns.
L.E. Gilbert homepage. Ecology and behaviour.
Briscoe Lab. Molecular evolution of eye pigment genes .
(visitado em 11/mar/2021)
https://www.scielo.br/scielo.php?script=sci_arttext&pid=S0101-81752006000300022 (três espécies e duas subespécies de Actinote)
https://comenius.susqu.edu/biol/202/dictionary%20of%20terms/a/apomorphic-character.htm (visitado em 15/3/2021)
https://www.techtudo.com.br/dicas-e-tutoriais/2018/05/oito-funcoes-do-powerpoint-que-pouca-gente-conhece.ghtml (visitado em 15/3/2021) Edição de ppt.
http://eaulas.usp.br/portal/video.action;jsessionid=D9BF562294DEC2910A2AB54522BE1944?idPlaylist=6214¤tPlaylistIndex=1 (aula sobre Cladística USP)
0 comentários:
Postar um comentário