O vitelo (do Latim: vitellus = gema do ovo) é uma bolsa revestida por membrana que acumula reserva. O saco vitelínico é responsável pelo armazenamento de nutrientes, como lipoproteínas, vitaminas e colesterol. É o primeiro anexo embrionário que surge e está presente em peixes, anfíbios, répteis, aves e mamíferos. Nos répteis e nas aves o saco vitelínico é bem desenvolvido. Enquanto que nos mamíferos é reduzido uma vez que a fecundação e o desenvolvimento é dentro do corpo da mãe, e a placenta nesse caso é responsável pela nutrição do embrião, e também pela produção de hemácias (eritrócitos = células vermelhas do sangue) nos primeiros estágios da vida.
ALANTÓIDE
É uma pequena bolsa ricamente vascularizada, encontrada apenas em répteis, aves e mamíferos monotremados (que colocam ovos = Equidna e Ornitorrinco). Origina-se de uma evaginação do intestino primitivo. Apresenta as seguintes funções: 1) em répteis e aves - armazenar os resíduos nitrogenados formados pelo embrião durante o desenvolvimento, 2) promover as trocas gasosas do embrião com o meio, e 3) promover a absorção do cálcio da casca do ovo para a formação do esqueleto. A descalcificação da casca facilita seu rompimento no momento da eclosão do filhote.
Nos mamíferos, isso não ocorre, pois o alantóide junta-se com o saco ou vesícula vitelínica, formando o cordão umbilical, que elimina as excretas nitrogenadas do sangue do embrião para o sangue da mãe através da placenta.
Nos mamíferos, os vasos sanguíneos do alantóide auxiliam na formação da placenta.
Os répteis, as aves e os mamíferos são amniotas e alantoidianos, por formarem âmnion e alantóide. Os peixes e os anfíbios são anamniotas e analantoidianos, por não formarem âmnion nem alantóide, pois seu desenvolvimento é na água e seus resíduos tóxicos podem ser liberados diretamente no meio ambiente.
âmnio (do Latin amnion: membrana ao redor do feto; e do Grego: ἀμνίον água corrente, pote onde o sangue das vitimas (animais) era coletado. O âmnio é uma bolsa ou cavidade em forma de saco repleta de líquido amniótico, onde o embrião permanece até o nascimento. O âmnio tem uma função protetora, permitindo que o embrião se desenvolva num ambiente úmido e evitando a perda de água pelo embrião. Além disso, o âmnio protege o embrião, amortecendo os choques mecânicos e aderência, alem disso, pode evitar variações de temperatura. Na evolução dos vertebrados os répteis foram os primeiros amniotas o que lhes permitiu a conquista definitiva do ambiente terrestre.
CÓRION
O cório ou córion (do Grego: χόριον) é a membrana extra-embrionária que existe durante a gravidez. Apresenta duas camadas: 1) camada externa chamada de trofoblasto (do grego: trophos de trephein: alimentar + blastos: germinar) que é formada por duas outras camadas (o citotrofoblasto e o sinciciotrofoblasto); e 2) uma camada interna formada pelo mesoderma somático. O córion é o revestimento externo do embrião e que confere proteção mecânica, proteção térmica e contra a entrada de microorganismos.
VITELO
O vitelo
(do Latim: vitellus, vitellum = gema do ovo) é uma bolsa revestida por membrana
que acumula reserva. O saco vitelínico é responsável pelo armazenamento
de nutrientes, como lipoproteínas, vitaminas, ácidos graxos e colesterol. É o primeiro
anexo embrionário que surge e está presente em peixes, anfíbios,
répteis, aves e mamíferos. Nos répteis e nas aves o saco vitelínico é
bem desenvolvido. Enquanto que nos mamíferos é reduzido uma vez que a
fecundação e o desenvolvimento é dentro do corpo da mãe, e a placenta
nesse caso é responsável pela nutrição do embrião, e também pela
produção de hemácias (eritrócitos = células vermelhas do sangue) nos
primeiros estágios da vida. Nos répteis e aves o vitelo (gema do ovo) contem aproximadamente 60 calorias (três vezes mais do que a clara). Um ovo grande, pode conter: 2,7 g de proteínas, 210 mg de colesterol, 0,61 g de carboidratos e 4,5 g de gordura total. Vitaminas A, D, E e K (solúveis na gordura), sendo um dos raros alimentos que contém vita D.
ALANTOIDE
É uma pequena bolsa em formato de salsicha (do grego: αλλαντοΐδα, allantoida, allantoides, pequena salsicha) ricamente
vascularizada, encontrada apenas em répteis, aves e mamíferos
monotremados (que colocam ovos = Equidna e Ornitorrinco). O alantóide Origina-se de
uma evaginação do intestino primitivo. Apresenta as seguintes funções: 1) em répteis e aves - armazenar os resíduos nitrogenados formados pelo embrião durante o desenvolvimento. 2) promover as trocas gasosas do embrião com o meio 3)
promover a absorção do cálcio da casca do ovo para a formação do
esqueleto. A descalcificação da casca facilita seu rompimento no momento
da eclosão do filhote. Nos demais mamíferos (marsupiais e placentários), isso não ocorre, pois o
alantoide junta-se com o saco ou vesícula vitelínica, formando o cordão
umbilical, que elimina as excretas nitrogenadas do sangue do embrião
para o sangue da mãe através da placenta. Nesses mamíferos, os vasos sanguíneos do alantoide auxiliam na formação da placenta. Os répteis, as aves e os mamíferos são amniotas e alantoidianos, por formarem âmnion e alantóide.
Os peixes e os anfíbios, por sua vez, são anamniotas (sem âmnio) e analantoidianos (sem alantoide), por não
formarem âmnion nem alantóide, pois seu desenvolvimento se dá na água e os resíduos tóxicos de seu desenvolvimento podem ser liberados diretamente no meio (água).
1- Acesso a alimentos no ar (insetos voadores), flores e frutos na copa de árvores ou localizar o alimento do de cima (roedores e peixes) e capturá-los sem que eles consigam escapar.
2- Maior eficiência e rapidez na procura de alimento e abrigo e fuga de predadores.
3- Migração para áreas mais favoráveis em busca de alimento e melhores condições climáticas (como ilhas, monhtanas...)
4- Dispersão a longa distância ou através de barreiras geográficas (povoar áreas antes inacessíveis ou difíceis por terra).
Requisitos para voar
1- Gerar com seus músculos ou a partir do ambiente uma força ascendente tal que contrabalanceie a força da gravidade.
2- Reduzir a resistência do meio, principalmente em vôos longos e rápidos.
3- Gerar força de impulso em várias velocidades e às vezes em espaços limitados.
4- Conservar a estabilidade, manobrar, frear e aterrissar de acordo com as necessidades.
5- Produção de força por um organismo reduzido.
6- Firmeza do tronco, costelas e asas durante o voo.
7- Produção e utilização eficiente da potência para o voo.
8- Possuir olhos adaptados e com boa acomodação visual (para manter o foco mesmo em velocidades elevadas).
Uma nova espécie de crocodiliano do Período Cretáceo (de cerca de 145,5 milhões a 65,5 milhões de anos atrás) foi descoberta no sudoeste da Tanzânia, com pernas mais finas e dentes que até então eram considerados exclusivos de mamíferos.
Os crocodilianos formam uma ordem de répteis aquáticos e ovíparos, que inclui os crocodilos, jacarés e o gavial, e são encontrados especialmente em regiões tropicais do mundo. Os crocodiliformes do Cretáceo, chamados de notossúquios, eram parentes distantes dos crocodilos e jacarés modernos.
Os notossúquios, que viviam nas massas terrestres do supercontinente Gondwana, tinham um nível de diversidade tanto morfológica como ecológica muito maior do que a encontrada nos atuais crocodilos. Um exemplo está na boca: em vez de fileiras de caninos cônicos e iguais, seus dentes eram divididos em tipos especializados em morder e em outros feitos para esmagar.
A descoberta da nova espécie, cujo fóssil foi encontrado em rochas de 105 milhões de anos, foi publicada na edição desta quinta-feira (5/8) da revista Nature.
Segundo Patrick O’Connor, do Ohio University College of Osteopathic Medicine, e colegas, na espécie, denominada Pakasuchus kapilimai, as fileiras superior e inferior de dentes entravam em contato de modo semelhante ao que até hoje havia sido observado apenas em mamíferos.
O animal também tinha o tamanho aproximado de um gato doméstico e era mais magro do que os crocodilianos atuais. Possuía ainda um pescoço flexível. Apesar das características inusitadas para a ordem, os cientistas afirmam que se tratava de um crocodiliforme.
A dentição mais complexa indica uma capacidade de processar alimentos que os crocodilos atuais – que simplesmente mordem e engolem – não possuem, mas os mamíferos sim.
De acordo com o estudo, o Pakasuchus kapilimai e outros notossúquios podem ter ocupado nichos ecológicos na Gondwana (supercontinente ao sul) que correspondiam aos preenchidos por mamíferos no hemisfério Norte.
“Um número de características dessa nova espécie – incluindo a redução no número de dentes e uma dentição especializada e similar à divisão em caninos, premolares e molares – é muito semelhante a características que foram críticas durante o curso da evolução dos mamíferos do Mesozóico para o Cenozóico”, disse O’Connor.
O artigo The evolution of mammal-like crocodyliforms in the Cretaceous Period of Gondwana Patrick M. O’Connor e outros, pode ser lido por assinantes da Nature em www.nature.com.
AS ADAPTAÇÕES PARA O VOO ESTÃO
RELACIONADAS A DUAS LINHAS PRINCIPAIS
1) FORMATO AERODINÂMICO
2) REDUÇÃO DO PESO CORPORAL
A maioria das aves esteja adaptada ao vôo, todavia existem algumas exceções. O pingüim, por exemplo, não voa, mas pode nadar e mergulhar. Já o avestruz pode caminhar e correr.
As aves surgiram durante a Era Mesozóica, cerca de 150 milhões de anos atrás. Acredita-se que elas tenham evoluído de répteis bípedes aparentados aos dinossauros. O registro mais antigo de uma ave é o fóssil da espécie Archaeopteryx lithografica. Embora o Archaeopteryx possuísse penas, ele também apresentava outras características, como uma longa cauda e ossos compactos, mais semelhantes aos répteis do que às aves atuais.
Adaptações para o vôo
As aves possuem diversas adaptações para o vôo que estão relacionadas ao formato aerodinâmico e à redução do peso do corpo. A presença de membros anteriores, transformados em asas, e de penas são algumas dessas adaptações. A pena é uma estrutura leve, mas ao mesmo tempo flexível e resistente. Além de atuar no vôo, é também um importante isolante térmico.
O isolamento térmico fornecido pelas penas foi essencial para o surgimento da endotermia nas aves. Isso permitiu que o calor produzido pela alta taxa metabólica desses animais não se dissipasse para o ambiente externo. Esse isolamento também protege as aves da perda de calor gerada pela passagem do ar pelo corpo durante o vôo.
Muitos dos ossos das aves são pneumáticos. Isso significa que o seu interior é oco, o que os torna mais leves. No interior dos ossos pneumáticos existem extensões do pulmão chamadas de sacos aéreos. Os sacos aéreos contribuem para a redução da densidade das aves, além de promoverem a refrigeração interna e atuarem nas trocas gasosas durante a respiração.
Outras características que contribuem para a redução do peso são: ausência de dentes, ausência de bexiga urinária e atrofia das gônadas fora da época reprodutiva. Além disso, as fêmeas geralmente só possuem um ovário.
O osso que une as costelas na região ventral, o esterno, apresenta uma projeção chamada de quilha. A quilha é o ponto de inserção dos fortes músculos peitorais, responsáveis pelo batimento das asas.
Digestão e excreção
A ausência de dentes impede que as aves triturem o alimento na boca, antes de engolir. Esta função é assumida pela moela, uma região do estômago cujas paredes são dotadas de músculos fortes. Na moela os alimentos são triturados e esmagados, ou seja, é realizada a digestão mecânica. Algumas espécies armazenam pedrinhas na moela, que aumentam o atrito e auxiliam na trituração do alimento.
Muitas espécies possuem um papo. O papo corresponde a uma dilatação da porção posterior do esôfago e serve para armazenar, temporariamente, o alimento coletado. Quando estão com filhotes, as aves podem armazenar alimento no papo para transportá-lo até o ninho e alimentar a prole.
As aves, assim como a maioria dos répteis, excretam ácido úrico, uma substância nitrogenada que é insolúvel em água. As excretas são eliminadas na forma de uma pasta branca junto com as fezes, que possuem coloração escura.
Reprodução
A fecundação das aves é interna e, assim como os répteis, elas possuem um ovo terrestre com uma casca protetora externa. Internamente, encontram-se os anexos embrionários (âmnio, cório, alantóide e saco vitelínico).
As aves são animais ovíparos, ou seja, botam ovos que completam seu desenvolvimento fora do corpo materno. Isso contribui para a redução do peso da fêmea, pois ela não carrega o ovo ou o embrião dentro de seu corpo, como na ovoviviparidade e na viviparidade.
As aves chocam os ovos e cuidam dos filhotes após o nascimento. Este comportamento de cuidado com a prole é chamado de cuidado parental. Em muitas espécies tanto a fêmea quanto o macho realizam esta atividade.
Órgãos dos sentidos
As aves possuem a visão e a audição bem desenvolvidas. Esses sentidos são essenciais para um deslocamento eficiente no ar, durante o vôo. Já o olfato é pouco desenvolvido na maioria das espécies.
A produção de sons é realizada através de uma estrutura situada na base da traquéia, a siringe. A vocalização possui uma grande importância na comunicação das aves, sendo uma característica particular de cada espécie. A vocalização surge para a informação do estado do individuo para chamar atenção do bando e dos filhotes e para a corte e cópula (reprodução).
1. Endotermia ou Homeotermia - o corpo apresenta temperatura constante (não varia com o ambiente). 2. Desenvolvimento de penas - isolamento térmico (forma um colchão de ar que isola o corpo do meio externo), auxiliam no voo e impedem que a água da chuva alcance a pele da ave. (As penas são impermeabilizadas com óleo proveniente da glândula uropigiana).
3. Desenvolvimento de ossos pneumáticos. Os ossos das aves desenvolveram câmaras internas que se comunicam com os sacos aéreos, tornando o esqueleto das aves mais leve, todavia muito resistente para suportar as fortes tensões geradas durante o voo.
4. Desenvolvimento de um sistema de sacos aéreos (diminuição do peso e da densidade) , facilitando alçar voo rapidamente, e aumentando a oxigenação do sangue.
6. Osso esterno se se expande para suportar grandes músculos peitorais, esse músculo gera enorme força durante o voo. O osso esterno adquire a forma de quilha (ou carena) para a inserção dos músculos peitorais adaptados ao voo, alem disso, auxilia na forma aerodinâmica da ave.
8. Desaparecimento da bexiga urinária, urina pastosa ou sólida (redução do peso).
9. Perdas dos dentes, mandíbula e maxilar transformado em bico (redução do peso) e auxílio na aerodinâmica.
10. Cuidado parental - os pais se revesam no cuidado com a prole, chocando os ovos e alimentando os filhotes até que estejam aptos a voarem e sobreviverem sozinhos no ambiente.
VOCE PODE QUERER DAR UMA LIDA NOS TEXTOS DESSES LINK:
1) Cite e explique 4 características apresentadas pelos répteis que possibilitaram a conquista do ambiente terrestre.
2) Construa uma tabela comparando peixes, anfíbios e répteis quanto a:
habitat, epitélio de revestimento, respiração, reprodução, membros locomotores, equilíbrio corporal, percepção do ambiente e excreção. (Use a internet)
3) Desenhe um ovo reptiliano e coloque o nome e a função em cada uma de seus constituintes (não esqueça dos anexos embrionários).
4) Porque os embriões dos répteis necessitam da albumina presente na clara do ovo e do vitelo do saco vitelínico?
Em referência ao seu modo de vida que no incio ocorre dentro d´água e depois os adultos são terrestres, embora não podendo se afastar da água, devido a sua reprodução e umidificação da pele, uma vez que esta não possui queratina.
ATIVIDADES
Resolver as questões do livro texto e trazer para aula.
OS CROSSOPTERIGIOS ERAM PEIXES COM NADADEIRAS LOBADAS E MUSCULOSAS QUE PODIAM SE ARRASTAR EM TERRA ENTRE UMA POÇA DE ÁGUA E OUTRA. OBSERVE OS OSSOS QUE SUSTENTAM AS NADADEIRAS ANTERIORES, QUE PASSARAM A FUNCIONAR COMO OSSOS DOS MEMBROS ADAPTADOS PARA ANDAR EM TERRA FIRME.
Um dos fósseis mais interessantes recuperados dos sedimentos Devonianos (com idade em torno de 375 milhões de anos), no hemisferio norte é o Tiktaalik roseae. Esse fóssil apresenta diversas características compartilhadas por todos os tetrápodas terrestres. Entres as características podemos encontrar:
1) um pescoço livre - que permite o movimento da cabeça, isto é, esses animais podiam mover a cabeça (virar a cabeça como nós). Esse movimento é muito útil na captura em ambientes confinados como ocorre nos pântanos. Além disso esses animais podiam se mover em água rasas o que deu a eles uma vantagem seletetiva frente aos peixes que não podiam se mover nessas águas.
2) Mobilidade na articulação do pulso - o que favorece a natação e o movimento em terra firme.
Tiktaalik roseae é a única espécie conhecida de Tiktaalik, um género de peixes sarcopterígeos (que possuem barbatanas com músculos) extintos do período Devoniano tardio (período da era Paleozóica compreendido aproximadamente entre 416 milhões e 359 milhões de anos atrás). O Tiktaalik apresenta muitas características típicas dos tetrápodas (animais de quatro membros locomotores (patas)). É um exemplo de várias linhas de sarcopterígeos antigos que desenvolveram adaptações aos habitats pobres em oxigénio das águas pouco profundas presentes no seu tempo, e que levaram à evolução dos primeiros anfíbios. Fósseis bem preservados foram encontrados na Ilha Ellesmere em Nunavut, Canadá.
Os primeiros fósseis do Tiktaalik roseae foram encontrados em 2004 no ártico do Canadá. Considerado um Fóssil de transição entre o Panderichthys e o Acanthostega, é conhecido e divulgado como Fóssil de transição que prova a transição dos peixes da água para a terra. O fóssil encontrado possui características comuns de peixes, como escamas e barbatanas; e de criaturas terrestres, como cabeça achatada, indício de pescoço, ombros, cotovelos e pulso.
ANFÍBIOS
CARACTERÍSTICAS GERAIS
1. Esqueleto - apresentam esqueleto ósseo formado por bicarbonato de cálcio e fosfato de cálcio.
2. Respiração - respiração pulmonar (a bexiga natatória dos peixes originou um pulmão primitivo que foi se desenvolvendo e se aprimorando ao longo do tempo).
Respiração cutânea - através da pele lisa, sem escamas, sem queratina e muito vascularizada (vasos sanguíneos que levam o sangue oxigenado para todo o corpo).
3. Dois pares de membros locomotores.
4. Pálpebras - por não estarem mais na água as pálpebras são essências para a umidificação dos olhos, sem elas os organismos ficariam cegos.
5. Coração formado por dois átrios e um ventrículo.
6. Primeiro grupo de organismos a apresentar tímpanos (membrana fina que capta sons por meio de ondas sonoras na atmosfera). Funções: captação de sons para proteção, localização de presas, localização de parceiros para cópula.
7. Primeiros organismos apresentar vocalização - funções: localizar coespecíficos para reprodução e demarcação de território e afastamento de competidores.
8. Sistema digestório completo terminando em uma cloaca.
TRANSIÇÃO PARA TERRA E EVOLUÇÃO DOS TETRÁPODAS
Peixes devonianos com nadadeiras lobadas e fortes que migraram para a terra lentamente através da seleção natural adaptaram-se a vida na interfácie terra-água, dando assim origem aos anfíbios.
Um dos fósseis mais interessantes recuperados dos sedimentos devonianos foi o Ichthyostega. Esse fóssil apresenta diversas características compartilhadas por todos os tetrápodas terrestres.
Entres as características podemos encontrar:
1) um pescoço livre - que permite o movimento da cabeça (dobrar o pescoço para os lados).
2) Mobilidade na articulação do pulso - o que favorece a natação e o movimento em terra firme.
Peixes devonianos com nadadeiras lobadas e fortes que migraram para a terra lentamente através da seleção natural adaptaram-se a vida na interfácie terra-água, dando assim origem aos anfíbios.
Moedas gregas de Éfeso e Metaponto cunhadas com artrópodes
(Moedas cunhadas na Grécia no seculo IV aC)
Os Artrópodes apareceram no registro fóssil durante o período Cambriano, compreendido entre 542 milhões e 488 milhões de anos atrás, aproximadamente, juntamente com muitos outros grupos de Invertebrados (como os grupos que já estudados anteriormente).
Mais de um milhão de espécies já foram descritas, a maioria delas são insetos.
De fato duas em cada três espécies conhecidas são artrópodes, e os membros do filo Arthropoda, podem ser encontrados em quase todos os habitats da biosfera.
Pelo menos um milhão das espécies animais descritas são insetos.
Uma citação de J.B.S. Haldane, um dos fundadores da síntese evolutiva, ilustra a concentração taxonômica da biodiversidade. Quando perguntado sobre o que ele poderia dizer sobre Deus observando a natureza, Haldane respondeu que ele devia ter "um desejo desordenado por besouros".
Eles são ecdizozoa, têm um esqueleto cuticular e, portanto, devem fazer a muda para crescer.
O filo mais bem-sucedido que eles consideram mais intimamente relacionado é o Nematoda. Como os nemátodos, eles necessitam trocar a cutícula à medida que crescem, mas a cutícula do artrópode é diferente, tanto na estrutura quanto na função, da dos nematoides. Os artrópodes possuem exoesqueletos segmentados rígidos. A muda tem um grande impacto na vida dos artrópodes; de acordo com alguns cálculos, os artrópodes passam 90% de sua vida se preparando para fazer a muda, mudando e se recuperando da muda mais recente.
Breve descrição
Arthropoda, de longe o maior e mais diverso táxon animal, inclui quelicerados, insetos, miriápodes e crustáceos, além de muitos táxons extintos, como por exemplo o Trilobitomorpha. O corpo segmentado possui um par de apêndices articulados em cada segmento. A epiderme secreta um exoesqueleto cuticular complexo que deve ser mudado para permitir aumento de tamanho. Os artrópodes existentes exibem especialização regional na estrutura e função de segmentos e apêndices, mas o ancestral provavelmente tinha apêndices semelhantes em todos os segmentos. O corpo é tipicamente dividido em cabeça e tronco, dos quais o tronco é frequentemente dividido em tórax e abdômen.
O intestino consiste no intestino anterior, médio e posterior e se estende por todo o comprimento do corpo da boca (anterior) ao ânus (posterior). O intestino anterior (estomodeu) e o intestino posterior (proctodeu) são invaginações epidérmicas, originados da ectoderme e são revestidos por cutícula, assim como todas as superfícies epidérmicas dos artrópodes.
O intestino médio é endodérmico e é responsável pela maior parte da secreção de enzimas, pela hidrólise e absorção dos nutrientes.
O celoma é reduzido a pequenos espaços associados às gônadas e sistema excretor.
A cavidade corporal funcional é uma hemocela espaçosa que é composta por um seio pericárdico dorsal e um seio perivisceral muito maior. Às vezes, há um pequeno seio perineural ventral ao redor do cordão nervoso ventral.
O sistema hemal inclui um coração dorsal, contrátil, tubular com óstios (um par por segmento) que bombeia sangue para a hemocela. Podendo ou não existirem aortas.
Os órgãos excretores variam de acordo com o táxon e incluem os túbulos malpighianos, nefrídios fechados internamente, que não se abrem para o exterior, são portanto, saculiformes, e glândulas coxais.
Os órgãos respiratórios também variam com o táxon e incluem as brânquias, pulmões foliáceos e traquéias.
O sistema nervoso consiste em um cérebro dorsal anterior constituído por dois ou três pares de gânglios, conectivos circum-entéricos e um cordão nervoso ventral com gânglios para cada segmento e nervos periféricos segmentares. Vários graus de tagmatização e cefalização são encontrados em diferentes taxa.
O desenvolvimento é derivado de ovos centrolécitos e clivagem superficial.
Existe frequentemente uma larva, embora o desenvolvimento seja direto em muitos. Os juvenis passam por uma série de ínstares (os estágios larvais) separados por mudas até atingir o tamanho adulto e a condição reprodutiva. Nesse momento, a muda e o crescimento podem cessar ou continuar, dependendo do táxon.
Características dos Arthropoda
1. Tagmose: o corpo esta segmentado, com segmentos, em geral, agrupados em duas ou três regiões distintas. Segmentos estes que se fundiram formando os tagmas. Assim, cada subfilo tem sua tagmose distinta. Por exemplo, a tagmose dos hexapoda é Cabeça, Tórax, Abdômen.
2. Apêndices articulados, pares e segmentados (a partir dos quais do filo recebeu o nome).
3. Simetria bilateral.
4. Um exoesqueleto quitinoso que é descartado na muda ou ecdise periodicamente e renovado na medida que o animal cresce.
5. Um canal alimentar tubular com boca e ânus (o sistema digestório completo).
6. Um sistema circulatório aberto, geralmente com um único vaso sanguíneo, consistindo uma estrutura tubular dorsal, com duas aberturas por segmentos (os ostíolos).
7. Uma cavidade corporal, o esquizoceloma, muito reduzido restito às gonadas e as vezes ao órgãos excretores.
8. A cavidade corporal na realidade é uma blastocele persistente, onde circula a hemolinfa.
9. O sistema nervoso consiste em um gânglio cerebroide anterior localizado acima do canal alimentar, um par de conetivos que parte do cérebro ao redor do canal alimentar, formando um anel nevoso circum-entérico e cordões nervosos ventral ganglionar, o primeiro gânglio é chamado de gânglio sub-entérico, e nervos sementares. Esse padrão é igual ao dos anelídeos.
10. Músculos esqueléticos estriados.
11. Respiração por meio de brânquias, pulões foliáceos, e traqueias que se abrem para o exterior por meio de espiráculo
12. Ausência de nefrídios
13. Dióicos com desenvolvimento direto ou indireto. Nos Hexapoda há uma diferenciação no desenvolvimento em relação a metamorfose, podendo ser ametábolo, hemimetábolo e holometábolo.
14. Triblásticos.
15. Protostômios. (Triplehorn and Jonnson, 2011).
Diversidade dos artrópodes em relação aos metazoários
Pelo critério da diversidade, distribuição e números absolutos, os artrópodes devem ser considerados como os mais bem sucedidos de todos os filos animais.
O sucesso dos artrópodes está relacionado a algumas características que são adaptações encontradas nesse filo:
EXOESQUELETO DE QUITINA
APÊNDICES ARTICULADOS
TAGMOSE OU TAGMATIZAÇÃO OU ARTROPODIZAÇÃO
ANNELIDA - ONYCHOPHORA - ARTHROPODA
Se os artrópodes são aparentados (relacionados) aos anelídeos ou se possuem um ancestral comum com este filo deveríamos encontrar um organismo muito parecido com um anelídeo e com um artrópode. Quem seria este elo intermediário entre artrópodes e anelídeos? Poeria existir tal organismo?
Sim esse organismo pode existir. Existe um organismo muito semelhante à um anelídeo composto por inúmeros segmentos mas já apresentando, em cada um desses segmentos um par de apêndices articulados (condição ancestral dos artrópdoes). Esse animal pertence ao filo Onychophora e chama-se Peripatus sp. Esse filo tem 12 gêneros e 76 espécies válidas (Costa, 2016).
Há uma espécie nativa do Brasil: Peripatus acacioi. Os registros fósseis indicam pouca mudança na organização básica nos últimos 500 milhões de anos.
Apresenta corpo alongado, entre 5 e 10 cm de comprimento, pele aveludada e numerosos pares de patas curtas e grossas, terminadas em pequenas garras afiadas.
Na cabeça há um par de antenas e um par de papilas secretoras de muco (jatos que imobilizam as presas). Vivem em ambientes muito particulares e úmidos (cutícula com pouca quitina), em florestas tropicais da África, Ásia, Austrália e América do Sul.
Onychophora
Semelhanças dos Onychophora com Annelida
As semelhanças entre onicóforos e anelídeos aparecem na organização muscular, em camadas sob a pele, no sistema excretor (nefrídios) e na estrutura dos órgãos reprodutivos.
As semelhanças com os artrópodes são o sistema circulatório aberto e o sistema respiratório traqueal, apendices articulados.
Peripatus spp
O filo Onychophora é um filo intermediário entre os anelídeos e os artrópodes (embora alguns pesquisadores consideram os onicóforos como sendo ancestrais dos uniramia = miriápoda (que incluem os quilópoda e diplópoda) e hexapoda = insecta, por possuírem apêndices unirramosos).
Os Onychophoros como o Peripatus sp ainda apresentam segmentação semelhante aos anelídeos, mas essa característica nos artrópodes (muitos segmentos corporais) reduziu-se ou pela perda dos segmentos ou pela fusão dos segmentos (tagmatização ou tagmose) ao longo da evolução.
Você já viu um bichinho com aparência aveludada, debaixo de um tronco podre ou andando sobre folhas em locais úmidos em um bosque?
A princípio, lembram uma minhoca devido à flexibilidade do corpo, que é formado por segmentos não muito evidentes. Locomovem-se por meio de pernas carnosas, que funcionam à base de musculatura, mas não possuem articulação. Apresentam unhas (ganchos) nas patas e um par de antenas. São carnívoros e, nas mandíbulas possuem dentes que servem para comer suas presas. Apresenta um aspecto aveludado, e podem ter vários tipos de coloração como cinza, verde-oliva, azul, alaranjada, preta e avermelhada, embora sua coloração típica é púrpura-escuro nas costas, com a parte ventral mais clara, em tom lilás-avermelhado. O seu tamanho varia, podendo medir até 10 cm. Os machos possuem entre 24 e 26 patas, enquanto que as fêmeas entre 26 e 28.
Apesar da aparência frágil, é um animal carnívoro e um grande predador. Para se alimentar de insetos e aranhas, ele envolve a presa com um jato de muco de endurecimento rápido, que imobiliza a vítima. Nas mandíbulas, possui dentes quitinosos que servem para comer as presas.
Conhecidos popularmente como onicóforos ou vermes veludo, esse grupo é um dos mais antigos que habitam a Terra, com fósseis datados de 450 a 500 milhões de anos. Os onicóforos sobreviveram às variações das condições atmosféricas, como a chuva, vento, umidade (elevada ou baixa) e a separação dos continentes, e tantos outros eventos, inclusive aquele que extinguiu os dinossauros. Ao longo de milhões de anos, não sofreram transformações significativas em seu design, mantendo sua forma original desde então. Por isso, são considerados por muitos “fósseis vivos” ou “elos perdidos”. Pertencentes ao filo Onychophora, são invertebrados de hábitos terrestres, esses animais são peças importantes no “quebra-cabeça” da evolução das espécies e dos artrópodes em particular. Fisiologicamente, o onicóforo apresenta sistemas que se assemelham aos de outros dois filos: anelídeos – ao qual pertencem minhocas e sanguessugas – e artrópodes – insetos, aranhas e crustáceos. Por esse motivo, desde que foi descoberto, o filo Onychophora é considerado um “elo perdido” entre eles.
Wieloch coordena pesquisas que buscam conhecer um pouco mais sobre os onicóforos, encontrado em parques e estações ecológicas de Minas Gerais em no Brasil todo. O trabalho já resultou, inclusive, na descrição de uma nova espécie. O pesquisador explica que, grupos de animais antigos que conservam uma forma muito semelhante aos registros fósseis, ajudam os pesquisadores a realizar um estudo da história evolutiva profunda desses organismo e, assim, estabelecer relações de parentesco – como, no caso do onicóforo, com os anelídeos ou artrópodes. “A importância do grupo não se limita ao estudo da evolução das espécies. Dentro de um contexto geral, os onicóforos são mais antigos até mesmo que os dinossauros e têm resistido, há mais de 400 milhões de anos, à todas as transformações que o nosso planeta passou”, completa o professor.
No mundo inteiro, são conhecidos 50 gêneros e 150 espécies diferentes do filo Onychophora, que podem ser encontrados em quatro continentes: África, Ásia, Oceania e América (Central e do Sul). Por isso, de acordo com o pesquisador, esses animais reforçam a teoria da Pangéia, na qual a Terra era apenas um único bloco que se dividiu e originou os atuais continentes. No Brasil, já foram coletados e foram identificados entre os seguintes gêneros de onicofóros Peripatus, Macroperipatus, Epiperipatus e Oreperipatus.
Para diferenciar cada um, os pesquisadores observaram características como coloração, o número de patas, e até pequenas saliências no corpo, chamadas papilas.
O Peripatus foi descoberto em 1955, e descrito por um casal de pesquisadores alemães, Eveline e Ernesto Marcus. Recebeu o nome científico de Macroperipatus acacioi em homenagem ao pesquisador que o descobriu, Dr. Acácio.
Em 1978, com o intuito de sua preservação, foi criada, no local, após mobilização do meio científico nacional e internacional, a Estação Ecológica do Tripuí. Essa foi a primeira unidade de conservação de um animal invertebrado no Brasil.
Além da Estação do Tripuí, o Peripatus, típico da região de Minas Gerais, já foi encontrado na Estação Ecológica de Peti, que fica entre os municípios de São Gonçalo do Rio Abaixo e Santa Bárbara, e no Parque do Ibitipoca, sudoeste do Estado. O primeiro contato do professor Wieloch com os onicóforos foi através do Macroperipatus acacioi, em 1998. Ele participou de um projeto apoiado pela FAPEMIG que tinha o objetivo de realizar, em conjunto com outros departamentos da UFMG, um estudo ecológico sobre a espécie. O Departamento de Genética, por exemplo, desenvolveu a parte da biologia molecular. “A nossa colaboração foi mais na parte de logística. Coletávamos os espécimes juntos e, depois da análise, os devolvíamos para a Estação”, conta.
A maior dificuldade da equipe é mesmo coletar os espécimes, pois os onicóforos são animais de hábitos noturnos que possuem uma distribuição populacional do tipo agregada. Assim, a busca pela espécie é totalmente casual. “O grupo não se distribui de maneira homogênea.
Espécie em extinção
A cada ano, o Ministério do Meio Ambiente (MMA) publica uma lista nacional das espécies da fauna brasileira ameaçadas de extinção. Na lista nacional, consta apenas o nome do Macroperipatus acacioi, classificado como “em perigo”. Segundo os pesquisadores, ao contrário da lista brasileira, a lista mundial já classifica todo o filo Onychophora como vulnerável. O problema é que existem poucos especialistas no Brasil. “A espécie é considerada vulnerável não só pelo aspecto de raridade, mas também porque sua susceptibilidade aos impactos ambientais, como queimadas e desmatamentos, é muito grande.”
A classificação dos Artrópodes é algo controversa. A maioria dos zoólogos concorda que existem provavelmente quatro linhas principais de evolução, que se acredita serem representadas pelos extintos Trilobitas, pelos Chelicerata (quelicerados) constituídos pelos caranguejo ferradura (Limulos sp., escorpiões, aranhas e piolhos), pelos Crustácea, constituídos as cracas, camarões, lagostas, caranguejos e tatuzinhos de jardim; e pelos Uniramia que inclui as centopéias, lacraias e insetos.
Contrastando com os grupos anteriores, que se pensa terem origem marinha, os Uniramia podem ter evoluído em terra. Algumas investigações recentes, parece indicar que pelo menos os Uniramia e provavelmente também os outros 3 grupos de Artrópodes podem ter tido uma origem separada de diferentes antepassados anelidianos e a evolução das características dos Artrópodes (artropodização) deve ter surgido, independentemente, pelo menos 2 ou até mesmo 4 vezes. Se assim for, Arthropoda deveria ser considerado como um superfilo e Trilobita, Chelicerata, Crustacea e Uniramia como filos. Tal interpretação não é unânime mas, no entanto, certos autores mantêm as quatro linhas evolutivas separadamente e atribuem-lhes a categoria de subfilos mantendo Arthropoda como filo.
Como sabemos pelo registro fóssil desde o pré-cambriano, os artrópodes constituem o mais abundante e diversificado grupo de metazoários.
Para o homem são importantes na medicina, como vetores de doenças, e na produção de anticoagulantes e analgésicos; na agricultura, eles são importantíssimos pois podem se converter em "pragas" agrícolas (gafanhotos) e ecologicamente sem os artrópodes praticamente não haveria polinização. Além disso, eles constituem fonte de alimento por ex: camarão, siri, lagosta etc.. Além de produtos de alto valor econômico como a seda, o mel e a cera de abelha.
A seguir são apresentados os filos do superfilo artropoda ou subfilos do filo artrópode.
SUBFILO TRILOBITOMORPHA
Os trilobitas são os mais conhecidos representantes do subfilo Trilobitomorpha.
Os Trilobitomorfa são organismos extintos e constituem um dos grupos de fósseis mais populares.
Eram artrópodes com um corpo trilobado, característica esta que dá origem à palavra trilobita.
Kainops invius Trilobitomorfa do Devoniano inferior (early devonian)
The trilobite body is divided into three major sections (tagmata):
1. cephalon; 2. thorax; 3. pygidium. Trilobites are so named for the three longitudinal lobes: 4. right pleural lobe; 5. axial lobe; 6. left pleural lobe (Trilobites are named for the three longitudinal lobes);
the antennae and legs are not shown in these diagrams.
The subdivisions can be further broken down into different areas used in describing trilobite cephalic morphology. 1. preocular area; 2. palpebral area;
Já foram identificados 20.000 espécies de trilobitas até o presente momento. Todos extintos durante o Paleozóico.
Esses organismos foram extremamente abundantes e constituíram a fauna dominante dos mares nos períodos Cambriano e Ordoviciciano, há cerca de 550 a 440 milhões de anos atrás.
Foram protagonistas da fauna marinha até ao episódio grande extinção em massa que ocorreu no final da era Paleozóica.
Pelo facto de terem sido organismos abundantes, amplamente distribuídos e constituídos por um exosqueleto rígido, deixaram-nos o seu testemunho através de um registo fóssil extremamente rico e bem documentado.
A maioria dos continentes atuais estavam cobertas de água durante o Paleozóico, razão pela qual os vestígios dos Trilobitomorpha fazem parte de muitas das rochas sedimentares marinhas dispersas pelo globo.
Apesar destes organismos serem maioritariamente marinhos, exploraram diversos habitats e estilos de vida. Grande parte eram organismos bentônicos, errantes percorrendo os fundos marinhos e as camadas superficiais dos sedimentos. Algumas destas espécies tinham apenas alguns centímetros de comprimento. Porém, algumas formas podiam alcançar entre 60 a 70 centímetros.
Muitos destes organismos que habitavam os fundos marinhos eram comedores de depósitos e escavadores. Algumas espécies bentônicas eram também predadores, alternando a captura direta de presas com a recolha de matéria orgânica em deposição.
SUBFILO: CHELICERATA OU QUELICERIFORMES
CARACTERÍSTICAS GERAIS DOS QUELICERIFORMES OU CHELICERATA
Chelicerata do latim, derivado do grego: χηλή = khēlē; chela: garra + κέρας, kéras = chifre, corno.
Os Cheliceriformes ou Chelicerata constitui uma das principais subdivisões do filo Arthropoda; esse subfilo inclui as classes Arachnida (aranhas, escorpiões, carrapatos, ácaros, entre outros), Pycnogonida e Xiphosura (límulo) (do grego ξίφος = xíphos, espada + οὐρά = ourá, cauda), bem como extintas classes Eurypterida (os escorpiões marinhos) para citar alguns exemplos.
Todos eles têm o corpo dividido em duas partes ou tagmas:
Prossoma ou cefalotórax, composto de seis segmentos, e é coberto por um escudo dorsal em forma de carapaça.
Opistossoma ou abdômen, com até 12 segmentos, e um télson pós-anal.
Em alguns grupos o opistossoma pode estar subdividido em duas partes o mesossoma e metassoma.
No prossoma, os quelicerados possuem seis pares de apêndices multiarticulados e unirremes são eles:
Um par de quelíceras;
Um par de pedipalpos;
Quatro pares de pernas locomotoras.
Ao contrário dos restantes artrópodes, os quelicerados não têm mandíbulas nem antenas, possuindo apenas olhos simples os ocelos.
Sua respiração é realizada por pulmão foliáceo e traqueias.
Crustáceos
São um grupo de animais mais comuns que conhecemos ex.: siris, caranguejos , tatuzinhos-de-jardim, lagostas, cracas e camarões.
Há mais de 67.000 espécies descritas de Crustáceos da fauna atual, e provavelmente um número 5 ou 10 vezes maior de espécies estão ainda para serem descobertas e catalogadas. Eles se apresentam como os animais mais abundantes, diversificados e com maior distribuição nos oceanos.
Os crustáceos exibem uma diversidade impressionante de forma, de hábitos e de tamanho. Os maiores são os caranguejos-aranha do Japão (Macrocheira kaempferi), com 4m de abertura de pernas, e o caranguejo gigante da Tasmânia (Pseudocarcinus gigas).
A maioria dos crustáceos são aquáticos, sendo encontrados em todas as profundidades dos diversos ambientes marinhos, planctônicos, bentônicos.
Também existem muitas espécies de água doce, além de grupos semi-terrestres (e.g. manguezais).
Alguns grupos obtiveram sucesso em ambiente terrestre.
Seu corpo está dividido ao menos em cabeça e tórax (e.g. Remipedia) e, na maioria das vezes, também em abdômen; possuem um número variável de pernas.
Uma característica distinguível dos crustáceos é a presença de dois pares de antenas. Além disso, eles podem possuir uma carapaça que, inclusive, nomeia o grupo.
Assim como todos como os outros artrópodes, apresentam um esqueleto externo (formado por cutícula) com impregnação de sais de cálcio e crescimento por mudas, além de apêndices articulados e corpo segmentado. Porém, diferentemente dos outros grupos, seus apêndices seguem um padrão birreme, isto é, composto por dois ramos. Apresentam desenvolvimento indireto e sua formal larval típica, é a náuplio (larva livre natante e com espinhos cefálicos).
Anatomia de um crustáceo terrestre e outros gêneros
Thermosbaenacea é uma ordem de pequenos crutáceos que medem entre 2,0 e 5,0 mm, que podem ser encontrados em ambientes extremos, como fontes termais de 45ºC, formações anquialinas e lencóis freaticos com alta concentração de sulfeto de hidrogênio, gás altamente tóxico para a grande maioria dos animais. O nome Thermosnbaenacea vem do grego (thermós = calor) e faz referência ao ambiente em que a espécie tipo (Thermosbaena mirabilis Monod, 1924) habita.
Os Crustáceos estão divididos em cinco subclasses e cerca de 30 ordens. A maioria das espécies de crustáceos são aquáticas e são encontradas especialmente em água salgada.
Classes
Remipedia
Cephalocarida
Branquiópoda
Maxillopoda
Malacostraca
Muitos crustáceos, incluindo lagostins, lagostas e camarões, são importantes como alimento humano. Os tatuzinhos de jardim (incluindo os pillbugs) pertencem à subclasse Malacostraca e à ordem Isopoda.
Os tatuzinhos são isópodes terrestres que são os únicos crustáceos de importância agrícola.
Os tatuzinhos de jardim são encontrados sob pedras, troncos e detritos no chão. Alimentam-se de matéria vegetal e podem se tornar pragas de plantas jovens. Os tatuzinhos de jardim se distinguem por um corpo deprimido, sete pares de patas e uma cabeça em forma de escudo: os outros segmentos do corpo se estendem para o lado. O primeiro par de antenas (antênulas) é vestigial; as antenas conspícuas representam o segundo par.
Os ovos são transportados pela fêmea em um saco com a ninhada na parte inferior do corpo. A respiração envolve brânquias emparelhadas na parte traseira inferior do corpo. Como as brânquias devem permanecer úmidas, os tatuzinhos de jardim não podem suportar a ambientes secos. Pillbugs são capazes de enrolar-se em uma bola apertada para proteção. Os crustáceos podem ser melhor mortos e preservados em 70-80% de ETOH.
Callinectes sapidus ou siri-azul com vista dorsal da fêmea (à esquerda) e vista ventral do macho (à direita).
Miriápodes:
Diplópoda e Quilópoda
Hexápodes
Hexapoda é o subfilo que compreende artrópodes com três pares de patas. Atualmente compreende as classes Entognatha e Insecta.
As ordens Zygentoma e Archaeognatha são de insetos sem asas (subclasse Apterygota) tendo como grupos irmãos os insetos alados, da subclasse Pterygota.
Os Hexapoda compartilham diversas características:
corpo dividido em três tagmas (cabeça, tórax e abdômen),
três pares de pernas torácicas,
um par de antenas,
três conjuntos de “peças bucais” (mandíbulas, maxilas e lábio).
O sistema de trocas gasosas primariamente por difusão, túbulos de Malpighi formados por evaginações do proctodeu (ou seja, de origem ectodérmica). Destas, a principal característica exclusiva (i.e. sinapomorfia) dos Hexapoda é a subdivisão do tórax em três segmentos, onde cada segmento porta um par de pernas locomotoras.
Structure and absorption spectra of the P. aristolochiae butterfly wing scales. (A) Image of a P. aristolochiae butterfly. (B) Microscopic image of the matt black region. A gradient of blackness is observed along each scale. (C) Cross-sectional SEM image of a matt black scale. (D) SEM images of a scale from the matt black region reveal that the air-filling fraction at the apex is higher (by about +59%) than at the base. The inset in the lower left corner shows the 2D Fourier power spectrum of the corresponding nanohole array. (E) Absorption spectra measured in the apex and base of a single scale from the matt black region of P. aristolochiae. The scale apex of a single scale exhibits higher absorption than the base.
Através de milhões de anos de evolução via seleção natural, a natureza criou materiais biológicos com várias propriedades funcionais para a sobrevivência. Muitas das arquiteturas naturais complexas, como conchas, ossos e favos de mel, foram estudadas e imitadas no design para fabricação de materiais com dureza e rigidez aprimoradas. Recentemente, mais e mais pesquisadores começaram a pesquisar as asas das borboletas, principalmente por causa de suas cores deslumbrantes. Verificou-se que a maioria dessas cores iridescentes são produzidas por estruturas fotônicas periódicas nas escamas que compõem as superfícies das asas. Esses materiais se tornaram recentemente um foco de pesquisa multidisciplinar em função de suas aplicações promissoras na exibição de cores estruturais e em sensores avançados, cristais fotônicos e células solares.
CARACTERÍSTICAS GERAIS DOS ARTRÓPODES
1. APÊNDICES ARTICULADOS
Do grego ἄρθρον, árthron: articulação, juntas + ποδός, podos: pé.
Apresentam pés ou patas ou apêndices articulados (apêndices com juntas móveis).
Ao longo da evolução os apêndices articulados se modificaram e se especializaram em funções como: locomoção (nadar, correr, saltar), alimentação, recepção sensorial, reprodução e defesa.
Apêndices articulados adaptados a diferentes funções.
2. PLANO CORPORAL DE UM ARTRÓPODE HIPOTÉTICO
Design de um artrópode hipotético
(traduza os nomes)
Trilobita
Filo Trilobitomorpha
Traça dos livros (ordem Thysanura)
Escamas do corpo das traças
CRUSTÁCEOS MARINHOS
CRUSTÁCEOS DE ÁGUA DOCE
3. TRIBLÁSTICOS
Apresentam três folhetos embrionários
Ectoderme
Mesoderme
Endoderme
4. SIMETRIA BILATERAL E TAMANHO
Milimétricos a grandes dimensões
Através do plano geral do corpo de um inseto podemos ver
O caranguejo aranha japonês Macrocheira kaenpferi;
Brachyuras: é o maior artrópoda vivo.
Maior espécie de lacraia viva atualmente
(Classe Quilópoda)
5. CELOMA
O amplo celoma (cavidade do corpo) do tipo esquizoceloma dos Anelídeos, importantíssimo para o movimento destes animais (esqueleto hidrostático), é vestigial nos Artrópodes adultos.
Também a musculatura, que nos Anelídeos esta organizada seguindo a forma geral do corpo cilíndrico com músculos circulares e longitudinais (estes percorrem todo o corpo no sentido longitudinal) que envolvem e dão forma ao corpo, se modifica nos Artrópodes, formando vários feixes que se fixam internamente ao exosqueleto (nos apódemas) e atuam, tal como nos apêndices, para elevar o corpo do solo, na água ou no ar, permitindo todo tipo de movimento.
6. TAGMATIZAÇÃO OU ARTROPODIZAÇÃO
Os Tagmas (do grego: τάγμα - tagma, plural tagmata - τάγματα) é um grupo especializado de múltiplos segmentos ou metâmeros coerentemente organizados em uma unidade morfológica funcional. Um bom exemplo é a tagmose de insetos: cabeça, tórax e abdômen. Asssim, pode-se dizer que a tagmatização ou artropodização é a fusão de segmentos com especialização.
Os ancestrais dos artrópodes apresentavam o corpo metamerizado (muito semelhante aos anelídeos), todavia, ao longo da evolução desse grupo ocorreu um processo de fusão desses metâmeros ou segmentos (unidades repetidas que formam o corpo) em grupos de segmentos especializados para realizar certas funções e anatomicamente diferenciados de outros segmentos, os tagmas do grego: ordem, hierarquia.
O processo evolucionário que criou os tagamata pela fusão e modificação dos metâmeros é chamado de artropodização, tagmatização ou tagmose, que é uma forma extrema de heteronomia mediada pelos genes Hox (Homeobox) e outros genes do desenvolvimento que os genes Hox influenciam.
Um caminho para a tagmose
Tagmose em Hexapoda
Mosca das frutas mostrando as zonas do corpo influenciadas por diferentes gentes Hox.
Assim, os artrópodes apresentam em consequência da tagmatização o corpo dividido em cabeça, tórax e abdome, em alguns grupos ocorreu uma fusão maior ainda e a cabeça funde-se com o tórax formando o cefalotórax. Desta forma, essas espécies apresentam cefalotórax e abdome. Há formas nos quais a fusão foi mais longe ainda fundindo o cefalotórax com o abdome, por exemplo nos ácaros e carrapatos, onde não se pode distinguir as partes do corpo. Ha apenas uma tênue linha que indica a separação do cefalotórax do abdome.
7. EXOESQUELETO
O exoesqueleto dos artrópodes serve como um local rígido de fixação dos músculos e como uma barreira contra a invasão microbiana e a perda de íons e água dos tecidos. No entanto, o exoesqueleto é rígido e não pode se expandir. Assim, quando um artrópode cresce até um determinado tamanho, deve periodicamente trocar sua cutícula externa (que constitui seu exoesqueleto esclerotizado), e uma nova cutícula se forma. A troca da cutícula antiga permite aumento no tamanho do corpo e outras alterações morfológicas. O processo geral pode ocorrer durante dias ou até semanas sem sinais externos óbvios, porque o novo exoesqueleto se forma sob o antigo. Somente quando a camada antiga está realmente sendo trocada, torna-se evidente que o artrópode está passando por uma fase de muda.
Seqüencialmente, a cutícula velha (agora chamada de exúvia) se abre e o artrópode sai. A nova cutícula (que consiste em uma camada epicuticular mais externa, uma exocutícula média e uma endocutícula interna) se expande e endurece por várias horas, principalmente como resultado da reticulação de proteínas e quitina (um polissacarídeo nitrogenado derivado da glicose) que constitui a quitina.
Vários hormônios esteróides regulam o processo de muda. Os três principais são hormônio juvenil, ecdisona (20-hidroxiecdisona) e bursicon. Em geral, o hormônio juvenil governa que tipo de cutícula e tecidos internos se formarão na próxima muda. Por exemplo, se o hormônio juvenil estiver presente durante um período crítico de um estágio larval, o próximo estágio será o larval. Se o hormônio juvenil estiver ausente, como ocorre no último estágio larval (ou estágio pupal em certos artrópodes), a próxima muda resultará em uma alteração para a forma adulta.
A ecdisona controla o próprio processo de muda e está presente antes da muda para transformar os tecidos para muda do exoesqueleto. Finalmente, o peptídeo neurohormônio bursicon, muitas vezes chamado de hormônio bronzeador, regula a expansão e o endurecimento da nova cutícula, bem como o escurecimento da cutícula para sua cor marrom/preta característica.
Após a muda, um artrópode é descrito como "teneral", um "inexperiente"; é "fresco", pálido e macio. Dentro de uma ou duas horas, a cutícula endurece e escurece após um processo de tanagem análogo à produção de couro. Durante esta curta fase, o animal se expande, uma vez que o crescimento é restringido pela rigidez do exoesqueleto. O crescimento dos membros e de outras partes normalmente cobertas pelo exoesqueleto rígido é obtido pela transferência de fluidos corporais das partes moles antes que a nova cutícula endureça. Uma aranha com um abdômen pequeno pode estar desnutrida, mas mais provavelmente sofreu ecdise recentemente. Alguns artrópodes, especialmente os grandes insetos com respiração traqueal, expandem seu novo exoesqueleto engolindo ou aspirando ar, já os artropodes aquaticos ingerem água. A maturação da estrutura e a coloração do novo exoesqueleto podem levar dias ou semanas em um inseto de vida longa; isso pode dificultar a identificação de um indivíduo se ele tiver passado recentemente por ecdise.
A ecdise permite que tecidos danificados e membros ausentes ou perdidos por lesões sejam regenerados ou reformados substancialmente. A regeneração completa pode exigir uma série de mudas, o coto vai se tornando um pouco maior a cada muda, até atingir um tamanho normal ou quase normal.
O aparecimento do exoesqueleto quitinoso rígido e a tagmatização ou tagmose ou artropodização são considerado como os principais fatores na irradiação adaptativa dos artrópodes. Imagine um animal como um anelídeo de corpo mole que passou a estar envolvido por uma armadura rígida composta de quitina. Nitidamente, essa armadura além de fornecer proteção contra predadores e variações ambientais, como temperatura e acidez do meio, perda de água, entre outros, levou ao aparecimento de mudanças relacionadas à locomoção e necessidades fisiológicas desses animais.
Desse modo, os artrópodes apresentam músculos estriados em feixes (e não mais organizados circularmente em cada metâmero e músculos longitudinais como nos anelídeos. Nos anelídeos os músculos circulares e longitudinais atuavam sobre uma cutícula flexível e sobre o celoma que promovia uma ação antagônica a esses músculos. Nos artrópodes músculos circulares e longitudinais seriam totalmente inefetivos pois o exoesqueleto não se deforma.
Assim, a musculatura dos artrópodes está organizada em feixes individualizados inseridos em apódemas que são projeções de quitina para dentro do exoesqueleto.
Estes músculos se prendem na parede interna do exoesqueleto em apódemas e vão até os apêndices (músculos extrínsecos) e aqueles que vão de um artículo a outro do apêndice (músculos intrínsecos).
Principles of insect morphology. Snodgrass, R. E. Ithaca, Cornel Univ. Press, 1993.
Feixes de músculos extrínsecos e intrínsecos
Todavia, o exoesqueleto dificulta o crescimento, surgindo a necessidade de o animal trocar ou mudar de exoesqueleto (ecdise) à medida que seu corpo aumenta de tamanho, durante o desenvolvimento. Durante a muda os artrópodes ficam mais vulneráveis à ação dos predadores, porém o enorme sucesso dos artrópodes como grupo revela que a artropodização foi um evento muito bem sucedido ao longo da evolução.
Comparação entre o corpo de artrópode e de um homem
Cefalotórax, abdômen e pós-abdômen em um escorpião
Relembrando: A quitina (C8H13O5N)n é um monossacarídeo derivado da glicose, constituído por muitas moléculas de N-acetilglicosamina (biopolímero). É uma amida entre glicosamina e ácido acético. Insolúvel em água, álcool, álcalis, ácidos diluídos, ou pelos sucos digestivos de outros animais. A quitina ocorre naturalmente em diversos organismos entre eles podemos citar: parede celular de certas bactérias, exceto arqueias, (como ácido N-acetilmurâmico) ligados com oligopeptídeos (o que constitui o peptídeoglicano); é o principal componente da parede celular dos fungos e também está presente na carapaça ou concha dos protoctistas foraminíferos, na cutícula dos nematóides, na rádula dos moluscos e no bico dos cefalópodes (polvos e lulas). Sendo altamente resistente, rígida e impermeável ela constitui o exoesqueleto dos artrópodes.
Estrutura do exoesqueleto e da cutícula de um lagosta
Modelo esquemático da cutícula da lagosta e do ambiente basal e apical, incluindo uma glândula dérmica com canal revestido de apatita com carbonato (2: 1–7: 1 Ca: P) (Kunkel & Jercinovic 2012) secretando um produto Xi / Xo. As trabéculas fosfáticas são 7: 1 Ca: P. Durante a produção de cutícula, Ca ^ 2 + e CO3 ^ 2- são importados para o espaço cutâneo do lado da hemolinfa, equilibrados pela expulsão de um próton. Na superfície externa da cutícula, a epicutícula regula uma lenta dissolução da camada de calcita, que produz uma corrente contínua de hidroxilos na camada não ondulada, aumentando seu pH à medida que o CaCO3 em dissolução se combina com a água. A camada não ondulada está finalmente em equilíbrio dinâmico com o oceano aberto, nominalmente em pH 8,2, mas provavelmente mais próximo de pH 7,8 no ambiente bentônico da lagosta. Uma lesão (seta com asterisco) disponibiliza carbonato de cálcio amorfo mais solúvel (ACC) para alcalinização rápida da camada superficial não ondulada.
Schematic model of lobster cuticle and basal and apical environment including a dermal gland with graded (2:1–7:1 Ca:P) carbonate apatite- lined canal (Kunkel & Jercinovic 2012) secreting a product Xi / Xo. Phosphatic trabeculae are 7:1 Ca:P. During cuticle production, Ca^2+ and CO3^2- are imported into the cuticle space from the hemolymph side, balanced by the expulsion of a proton. At the cuticle outer surface, the epicuticle regulates a slow dissolution of the calcite layer, which produces a continuous stream of hydroxyls into the unstirred layer, raising its pH as the dissolving CaCO3 combines with water. The unstirred layer is ultimately in a dynamic equilibrium with the open ocean, which is nominally at pH 8.2, but probably closer to pH 7.8 in the lobster's benthic environment. A lesion (asterisk arrow) makes more soluble amorphous calcium carbonate (ACC) available for rapid alkalinization of the superficial unstirred layer.
Cutícula
Esquema do integumento de um artrópode
(Storer & Usinguer, 1982)
(Fontes das figuras: Triplehorn & Johnson, 2011, Ruppert & Barnes 1995; notas de aula de Morfologia I e II, 1995).
The molecular organization of the epicuticle (the outermost layer) of insect wings is vital in the formation of the nanoscale surface patterns that are responsible for bestowing remarkable functional properties. (Dragonfly Hemianax papuensis) Using a combination of spectroscopic and chromatographic techniques, including Synchrotron-sourced Fourier-transform infrared microspectrometer. (Researche Gate).
MUDA OU ECDISE
Como já foi salientando a presença do exoesqueleto quitinoso exige um constante processo de ecdises (mudas). Mas como se dá mesmo este processo? Apesar de todos os artrópodes sofrerem mudas, na maioria das vezes a muda está vinculada ao desenvolvimento e ao crescimento. Em alguns artrópodes por ex., em crustáceos, quelicerados, a ecdise continua acontecendo mesmo que o animal atinja a fase adulta de seu ciclo de vida. Até que o animal passe pela última muda, o processo de ecdise é praticamente contínuo e o animal pode passar 90% de sua vida se preparando, passando ou concluindo uma fase de muda.
Os estágios entre mudas são chamados de instares (instar ovo, instar larva etc) e é durante esse estágio de intermuda que o crescimento real e efetivo dos tecidos ocorre, mesmo que não ocorra o crescimento no tamanho externo.
O exoesqueleto só é trocado quando todo espaço interno já foi preenchido. Podemos dividir todo esse processo em algumas etapas: intermuda; pré-muda ou pré-ecdise; muda; e pós-muda ou pós-ecdise. (Brusca & Brusca (2007), Ruppert et al. (2005) e Hickman et al. (2004)
Durante a intermuda o animal realiza suas atividades normais, se alimenta, acumula reservas para uma próxima muda e, se for maduro sexualmente, pode se reproduzir.
A pré-muda por sua vez é um estágio dito preparatório, onde ocorrem mudanças significativas: o animal entra neste estágio a partir da fase de intermuda e sai dele no momento em que a cutícula velha se descola do animal, mas ainda o está cobrindo, e a cutícula nova está parcialmente formada; durante este estágio o animal não se alimenta. Na pré-muda a epiderme secreta a nova epicutícula e libera o líquido exuvial ou de muda (que contém enzimas) entre a nova epicutícula e a cutícula velha (é bom lembrar que a cutícula é subdividida em epicutícula camada protéica fina impregnada de lipídios e procutícula ou corpo da cutícula se divide em exocutícula, endocutícula, camada calcária e camada membranosa). A liberação do liquido ou fluido exuvial que contem enzimas provoca a dissolução da velha endocutícula, e os produtos solúveis são reabsorvidos e armazenados no interior do corpo. Uma nova exocutícula é secretada enquanto a velha endocutícula está sendo reabsorvida.
Nesse momento o animal está coberto por dois exoesqueletos, ambos formados apenas por epicutícula e exocutícula, já que a nova endocutícula ainda não foi secretada e a endocutícula velha já foi destruída.
Ecdise em artrópode. (A) Exoesqueleto na íntegra e epiderme subjacente durante a intermuda. (B) Separação da epiderme e secreção do líquido de muda e de nova epicutícula durante o início da pré-muda. (C) Degradação da endocutícula velha e secreção da procutícula nova. (D) Final da pré-muda: dois esqueletos - o velho e o novo. (adaptado de Ruppert et al., 2005).
Muda ou ecdise em uma borboleta Dryas iulia (Fabr., 1779)
O estágio mais curto é a muda propriamente dita. Nesse momento, o animal rompe o exoesqueleto velho, geralmente em um lugar de articulação localizado nas linhas de fratura, normalmente no dorso, onde a epicutícula é fina. nesse momento o animal fica vulnerável já que o exoesqueleto novo ainda não endureceu. Pois ainda não ocorreu a tanagem da exocutícula. Nessa fase o animal geralmente não é avistado, pois se protege em lugares abrigados. O animal absorve água (se for aquático) ou ar (se for terrestre) através do intestino e aumenta o volume sanguíneo e a cutícula se rompe por diferença de pressão; o animal finalmente se livra do velho exoesqueleto.
A cutícula nova precisa agora ser esticada para poder acomodar o crescimento dos futuros tecidos. Na última fase, pós-ecdise, a nova endocutícula é secretada e a cutícula como um todo é endurecida através de esclerotização ou redeposição de sais de cálcio (nos crustáceos).
Como discutido anteriormente, a maioria dos artrópodes realiza o processo de muda com objetivo de crescimento. Para esses animais, quando se atinge a forma adulta, entra-se no estágio de não-ecdise, e permanecerá assim pelo tempo em que durar sua vida, pois chegou ao instar final.
Nos insetos, a muda também acontece na fase da metamorfose que levará o animal ao estágio de imago ou adulto.
Todo o processo de muda é regulado pelo hormônio da muda ou ecdisona e obriga o animal a crescer, aparentemente, aos saltos, de forma descontínua.
A libélula tem 300 milhões de anos de evolução ao seu lado. A empresa Animal Dynamics passou quatro anos escrevendo um software que opera o drone lançado à mão como um inseto e permite pairar em rajadas de mais de 20 nós (23 mph ou 37 km / h). De 22 a 27 nós é classificado como uma "brisa forte". (BBC)
O exoesqueleto dos artrópodes é formado por cutícula em alguns casos ela é espessa e com deposição de minerais como cálcio, como nos crustáceos.
FUNÇÕES DO EXOESQUELETO
Entre as funções importantes do exoesqueleto dos artrópodes as principais são: Sustentação, suporte para os músculos e proteção.
Sustentação
através dos apêndices articulados o peso do animal é transferido para o substrato, e assim ele pode: andar, correr, saltar, nadar e voar.
Suporte para os músculos
É sobre o exoesqueleto que os músculos atuam, fornecendo uma força antagônica para que o movimento aconteça. Assim, os músculos estão inseridos em apódemas que são projeções do exoesqueleto para dentro do corpo.
Proteção O exoesqueleto funciona como uma armadura contra os predadores e ainda auxilia evitando a perda de água (desidratação).
Revestimento
proteção física (mecânica) contra ataques de predadores
Proteção contra desidratação (perda de água)
Proteção em geral: cutícula quitinosa espessa e biomineralizada (crustáceos)
Torna o exoesqueleto defensivo mais resistente, o que permite o animal crescer mais e ficar mais forte (por possuir um exoesqueleto mais rígido).
Comunicação Cores vistosas = coloração de aviso, ou camuflagem com o objetivo de se confundir com o meio ambiente.
Função mecânica Os segmentos são articulados possibilitando todo tipo de movimento (tanto dos apêndices como do corpo), além disso, os apêndices articulados transferem o peso do artrópode para a superfície onde ele se encontra possibilitando uma vasta gama de atividades.
8. SISTEMA CIRCULATÓRIO
Sistema circulatório aberto
9. SISTEMA DIGESTÓRIO
1) Originalmente um tubo simples.
2) Uma única camada de células epiteliais.
3) Intestino anterior (estomodeu): invaginação da ectoderma.
4) Intestino médio (mesêntero): formado pela endoderma.
5) Intestino posterior (proctodeu): invaginação da ectoderma. (Leite, 2011)
Funções básicas
1) Intestino anterior: estocagem.
2) Intestino médio: enzimas digestivas e absorção.
3) Intestino posterior: coleta, condução e eliminação de excretas, reabsorção de água e sais. (Leite, 2011)
Intestino anterior
1) Cutícula presente chamada intima: previne desvio de alimento para o interior do corpo e ajuda na
estocagem.
2) Cavidade pré-oral: manipulação e ingestão do alimento.
* proventrículo ou moela: redução do alimento, separação do pólen do néctar
em abelhas (o pólen vai para o intestino médio para a digestão) e o néctar é
regorgitado dando origem ao mel) e seleciona o alimento para digestão por meio da
válvula estomodeo (ou esofagiana ou cardíaca) presente na junção do intestino
anterior e mediano.(Leite, 2011)
Intestino médio
1) Ausência de intima.
2) Da dobra da válvula cardíaca até antes da inserção dos túbulos de Malpighian.
3) Ocorre a digestão e absorção do alimento para a cavidade do corpo.
4) Cecos gástricos: evaginações que formam sacos, onde secretam enzimas digestivas também.
5) Ventrículo ou estomago
* células diferenciadas da epiderme para secreção, absorção e células regenerativas.
* membrana peritrófica: secretada pela epiderme (tecido não vivo), envolve o alimento, protege as células do epitélio da abrasão, protege o corpo de invasão de parasitos, separa o ventrículo em duas partes (ectoperitrófico e endoperitrófico). * Modificações: câmaras de filtro em insetos sugadores de seiva, onde ocorre rápida eliminação de excesso de água e sais para concentração dos nutrientes. Problema maior com insetos sugadores de seiva do xilema (pequena quantidade de aminoácidose alta concentração de ácidos orgânicos) do que os que sugam seiva do floema (alta concentração de açúcar e aminoácidos).
6) A câmara de filtro permite que o excesso de água passe diretamente da região final anterior do intestino médio ou da região final do posterior do intestino médio ou túbulos de Malpighian, diretamente para o intestino posterior.(Leite, 2011)
Divisão do sistema digestório (Gallo et all, 2002) apud (Leite, 2011)
Intestino posterior
1) Entrada do intestino posterior: válvula pilórica.
2) Não ocorre digestão.
3) Transporte de alimento não digerido para fora
4) Balanço de sais e água.
5) Intima mais fina que o do intestino anterior permitindo limitada absorção de pequenas moléculas tais como água, sais e aminoácidos removidos do sangue pêlos túbulos de Malpighi. (Leite, 2011)
Digestão e absorção de alimentos
1) Envolve uma série de ações enzimáticas.
2) Carboidratos: absorvidos como monossacarídeos (enzima: amilase, entre outras).
3) Maioria dos insetos não produzem celulase, não sendo capazes de digerir a celulose:
* Simbiontes intestinais.
* Térmitas: perdem os simbiontes em cada muda, mas obtém por meio de trofaloxia (troca mutua de material alimentar parcialmente digeridos).
4) Lipídeos: ingeridos geralmente como triglicerídeos, são absorvidos como diglicerídeos, monoglicerídeos ou ácidos graxos livres (enzimas: lípases, esterases).
5) Proteínas: absorvidas como polipeptídeos, dipeptídeos e aminoacidos (enzimas: proteases).
Absorção
1) Passiva: difusão osmótica por diferença de concentração entre lúmen do intestino médio e a hemolinfa.
2) Ativa: processo de movimento de substâncias contra um gradiente de concentração com gastos de energia. Ex.: o transporte ativo de sódio tem um papel importante na difusão da água para dentro e para fora do epitélio celular do intestino médio influenciando a absorção passiva de nutrientes por meio da parede do intestino. (Leite, 2011)
Glândulas salivares
1) Umedecer e amaciar o alimento.
2) Solvente para açúcar.
3) Meio de enzimas digestivas
Amilases atuam sobre carboidratos.
Invertases atuam sobre carboidratos.
Pectinases atuam sobre a pectina, presente nas parede das células vegetais.
Hialuronidase, em predadores: para dissolver conteúdo interno das presas).
3) Lipídeos: energia química, hormônios, estoque de energia e colesterol para crescimento e desenvolvimento.
4) Vitaminas: especificas funções fisiológicas. Ex.: vitamina A, visão.
5) Minerais: para crescimento e desenvolvimento normal. Ex.: K, P, Mg, Na. Ca. Cu e Zn.
6) Água.
7) Microorganismos: levedura, fungos, bactérias e protozoários (simbiontes)(Leite, 2011).
10. TROCAS GASOSAS
Diagrama esquemático das amostras traqueais utilizadas para MEV e microscopia óptica. (a) Localização das principais traquéias torácicas dentro da barata americana. (b) Traquéias torácicas dorsais principais esquerda e direita, conectadas por uma traquéia secundária (ST). (c) Espécimes em anel, que são aproximadamente cilindros elípticos, foram cortados ao longo do eixo longitudinal para produzir dois espécimes em meio anel, cada um com a maior curvatura na região central indicada em cinza. (Estrutura das traqueias de barata).
Respiração traqueal em barata americana, hexápoda (Fonte Researche Gate).
11. SISTEMA EXCRETOR
Os órgãos renais de insetos geralmente exibem altas taxas de transporte de ânions inorgânicos e orgânicos e, portanto, fornecem modelos úteis para o estudo do transporte de ânions epiteliais e seu controle. Túbulos de Malpighi isolados de algumas espécies secretam um volume de líquido iso-osmótico igual ao seu próprio volume em 10 a 15 segundos, o que significa que o conteúdo de Cl celular é trocado a cada 3 a 5 s.
O transporte de ânions também pode ser alcançado contra gradientes termodinâmicos extremos. A concentração de K + e Cl- no lúmen dos túbulos Malpighianos de alguns besouros do deserto se aproxima ou excede a saturação. Um cotransportador basolateral de Na +: K +: 2Cl desempenha um papel importante no transporte de íons vetoriais nos túbulos de Malpighi de muitas espécies, mas também há evidências de acoplamento do transporte de Cl ao movimento de uma única espécie catiônica (Na+ ou K+).
Embora uma H+ -ATPase vacuolar apical tenha um papel primário na energização da secreção transepitelial de cloreto por canais ou cotransportadores no segmento secretor do túbulo de Malpighi, várias ATPases diferentes foram implicadas na reabsorção de Cl- pelo túbulo ou intestino inferior dos túbulos de Malpighi. Sabe-se que o transporte de cloretos é controlado por vários neuropeptídeos, aminas e segundos mensageiros intracelulares. Os epitélios renais dos insetos também são importantes na excreção de ânions orgânicos potencialmente tóxicos, e os transportadores envolvidos podem desempenhar um papel na resistência a inseticidas de origem natural ou antropogênica.
DESENVOLVIMENTO
Quando comparamos o desenvolvimento dos artrópodes com os demais metazoários notamos que eles são radicalmente diferentes. E porque essa diferença? Essa diferença se dá devido a possuírem um esqueleto esterno rígido.
Assim, quando observamos seu desenvolvimento notamos que eles podem passar por diferentes fases em sua vida, iniciando obviamente com a fecundação temos o OVO, deste, eclode uma larva de primeiro instar, que ao se alimentar continuamente cresce e necessita mudar seu exoesqueleto. Desta forma, em um determinado momento ela para de comer, encontra um local protegido e troca seu exoesqueleto a essa muda do exoesqueleto antigo por um novo chamamos de muda ou ecdise. E assim sucessivamente ela sofre várias mudas em estágio larval, (de larva I, II, III, IV, V) e deste último estágio ela passa então por um período chamado pupa (onde todos os órgãos larvais são destruídos) e novos órgãos são construídos, transformando-se em em um organismo adulto ou imago. (holometábolos).
Todavia nem todos os artrópodes passam por todos esses estágios exatamente da mesma forma; em alguns do ovo já eclode um individuo semelhante ao adulto exceto que falta as asas (características do adulto) (hemimetábolos) e em outros casos do ovo sai um indivíduo pequeno mas com todas as características do adulto (ametábolos).
Como consequência da rigidez do exoesqueleto e do crescimento, todos os artrópodes devem realizar uma ou mais trocas do exoesqueleto ao longo de sua vida a medida que o animal cresce. Essa troca do exoesqueleto "velho" e apertado por um novo chama-se muda ou ecdise.
QUANTO AO DESENVOLVIMENTO: AMETÁBOLOS, HEMIMETÁBOLOS E HOLOMETÁBOLOS
Visão diagramática das atividades de mudança da epiderme durante o quarto e o quinto ínstares larvais e o desenvolvimento da pré-pupa (phareate pupal) no verme do tabaco, Manduca sexta (Lepidoptera: Sphingidae) em relação ao ambiente hormonal. Os pontos nas células epidérmicas representam grânulos do pigmento azul insecticianina. ETH, hormônio desencadeador de ecdise; EH, hormônio de eclosão; JH, hormônio juvenil; EPI, EXO, ENDO, deposição de epicutícula pupal, exocutícula e endocutícula, respectivamente. Os números no eixo x representam dias. (After de Riddiford 1991).
HOLOMETÁBOLOS OU METAMORFOSE COMPLETA
Desenvolvimento de uma mosca (HOLOMETABOLIA)
Devido ao crescimento do organismo e o não crescimento do exoesqueleto os artrópodes passam por eventos chamados mudas. Nesse caso esta sendo representado o desenvolvimento de uma mosca doméstica (Musca domestica), que ao longo de seu desenvolvimento de ovo até o estágio adulto passa por diferentes instares (ovo - larva - pupa - imago ou adulto). Nesse caso dizemos que esse organismo apresenta metamorfose do tipo holometamorfose ou metamorfose completa; exemplos: mosca doméstica, besouros, borboletas, mariposas.
Estágios da metamorfose (holometabolia) completa de um besouro
Estágios da metamorfose (holometabolia) completa de uma mariposa
Ovo de uma espécie de borboleta chamada
Heliconius charithonia
Estudo dos estágios imaturos (ovo) de Dryas iulia Fabr. 1775
Desenvolvimento em uma borboleta Ithominae. Os artrópodes apresentam crescimento descontínuo quando comparados com os demais metazoa, pois enquanto todos os demais animais apresentam um crescimento contínuo e gradual os artrópodes apresentam um crescimento aos saltos ou descontínuo.
E isso se deve ao fato de que eles devem trocar o exoesqueleto de tempos em tempos, pois ao se alimentarem as células se dividem por mitose enquanto o exoesqueleto rígido limita esse crescimento.
Durante o desenvolvimento larval, alguns insetos são capazes de regenerar seus apêndices após sua perda acidental. Se a perda ocorrer no início de um estágio de desenvolvimento, antes da produção do hormônio da muda, o apêndice será reformado na próxima muda. A regeneração ocorre nas formas larvais de Blattodea, Phasmatodea, em alguns hemípteros, ortópteros e insetos holometábolos. A regeneração das estruturas cuticulares só pode ocorrer na muda, pois é o único momento em que a nova cutícula é produzida. Consequentemente, a regeneração de apêndices não ocorre em adultos e é restrita apenas aos estágios larvares. A regeneração dos músculos e partes do sistema nervoso também ocorre durante os estágios de desenvolvimento.
Comente esses gráficos sobre o crescimento dos artrópodes
HEMIMETÁBOLOS OU METAMORFOSE INCOMPLETA
Ninfa
AMETÁBOLOS OU SEM METAMORFOSE
A traça dos livros, é um artrópode ametábolo; do ovo eclode um indivíduo igual ao adulto apenas em tamanho menor. Para chegar ao tamanho adulto a traça passa por várias mudas do exoesqueleto.
Cavidade do corpo é agora um hemocelo (cavidade onde circula o sangue) por isso dizemos que os artrópodes apresentam sistema circulatório aberto.
Ausência de repetição de órgãos como nos anelídeos.
Setas (cerdas) são substituídas por apêndices articulados
Uma possivel explicação para o surgimento dos artrópodes é a existência de um grupo intermediário entre os anelídeos e os artrópdas como mostrado no esquema abaixo.
(Snodgrass, R.E. Principles of insect morphology)
Assim, podemos entender que de um ancestral "vermiforme" como os anelídeos apareceram os onicóforos e de um ancestral semelhante a um onicóforo surgiram os artrópodes, através de uma grande irradiação adaptativa, para ocupar todos os ambientes existentes. Assim, vamos encontrar artrópodes em todo tipo de habitat (terra, água e ar).
Cladograma mostrando as características de cada taxa dos filo artrópoda. Note que os diplópoda (milípedes) e os quilópoda (centípedes) eram agrupados numa antiga denominação (para alguns subfilo para outros superclasse) chamada miriápoda (Myriapoda milhares de pés; por apresentarem muitos apêndices articulados locomotores).
(Definição de miriápoda do "A dictionary of entomology. Gordh & Headrick)
Cladograma mostrando as relações entre os subfilos dos artrópodes.
Os Atelocerata são os artrópodes mais derivados e se dividem em muitas ordens, cada uma delas adaptada a um nicho distinto.
COMPARAÇÃO ENTRE ANELÍDEOS E ARTRÓPODES
Triblásticos
Três folhetos embrionários (Ectoderme, Mesoderme e Endoderme)
Protostômios
O blastóporo origina a boca
Ectoderme
São considerados componentes ectodermais do tubo digestório: o estomodeo, o proctodeo e os túbulos de malpighi (excreção), uma vez que estes componentes apresentam caráter epitelial assim como a epiderme, e nunca demonstram características mesenquimais (Reuter, 1994). Assim, a ectoderme forma a epiderme que secreta a cutícula, o intestino anterior, o proctodeo, do embrião, o intestino posterior, o estomodeo, e os túbulos de Malpighi.
Cutícula
A cutícula nos anelídeos é fina e iridescente, nos artrópodes é espessa com no mínimo três camadas (epicutícula, exocutícula e endocutícula) e pode sofrer deposição de carbonato de cálcio nos crustáceos ou cera nos traqueata.
Mesoderme
A mesoderme é responsável pelo revestimento da cavidade do corpo, músculos, vasos sangüíneos e gônadas.
Endoderme
O tubo digestório, como já visto em D. melanogaster é formado por uma porção ectodermal e uma porção endodermal. A endoderme propriamente dita é responsável pela formação da porção mediana do tubo digestório.
Arquétipo de artrópode ancestral mostrando a origem de algumas estruturas
Metameria
Corpo segmentado, metâmeros são a unidade de desenvolvimento e expressão genética. Nos artrópodes a metameria heterônoma, que é a divisão do corpo em tagmas. A tagmose ou artropodização, que é a fusão dos metâmeros para formar os tagmas cabeça, tórax e abdômen.
Septos
Nos anelídeos os septos dividem cada segmento do corpo e consequentemente a cavidade do corpo o celoma esquizocélico. Já nos artrópodes os setos desapareceram e ha uma grande cavidade do corpo, a hemocela perivisceral, preenchida pelo sangue.
Celoma
Celoma esquizocélico em ambos, nos anelídeos o celoma em cada metâmero esta dividido em direito e esquerdo separados pelo peritônio que forma os septos; enquanto que o celoma dos artrópodes é muito reduzido e restrito às gonadas e aos nefrídios, ou perdido e o que resta é uma blastocele persistente com funções circulatórias a hemocele.
Cerdas ou setas
Pares de setas ou cerdas, arranjadas metamericamente (repetidas) com apêndices não-articulados, os parapódios nos poliquetos, que auxiliam no deslocamento e nas trocas gasosas (brânquias). Nos artrópodes há apêndices articulados para executar as mais variadas funções.
Músculos
Nos anelídeos ocorrem músculos circulares e longitudinais repetidos em todos os segmentos e bem desenvolvidos. Nos anelídeos é impossível que isso ocorra pois o exoesqueleto é rígido e não se deforma. Então ocorrem músculos organizados em feixes individualizados e está dividida em músculos intrínsecos (ligam artículos entre si) e extrínsecos (que ligam o corpo ao artículo do apêndice).
Sistema nervoso
Bem desenvolvido composto por um par de gânglios cerebrais dorsal semelhante a um cérebro que se localiza acima e à frente da faringe e um anel de cordões ao redor da faringe que conecta-se a um gânglio subfaringeal, a partir do qual um par de cordões nervosos fundidos corre posteriormente ao logo de todo o corpo com um gânglio nervoso para cada segmento. Já nos artrópodes o sistema nervoso é bem semelhante ao dos anelídeos no padrão geral (com um gânglio cerebral dorsal e um anel nervoso circumentérico e um cordão nervoso ventral ganglionar e o primeiro gânglio é chamado de gânglio subentérico e nervos segmentares para cada segmento do corpo.
1. Os artrópodes são o grupo mais numeroso de espécies dentro do reino Metazoa. Eles muito provavelmente evoluíram de um ancestral comum com os anelídeos e moluscos.
2. são organismos triblásticos, com celoma esquizocélico reduzido, e a cavidade do corpo é uma blastocele persistente que é a hemocele.
3. Na evolução dos artrópodes a cutícula ou o exoesqueleto de quitina constituiu-se como desenvolvimento central permitiu o grande sucesso alcançado por esse grupo de metazoa. Além disso, a divisão do esqueleto em placas (tergitos, esternito e pleuritos), e os apêndices articulados, tornou o movimento possível e sofisticado e mudas periódicas do exoesqueleto regulados pelo hormônio ecdisona permitiu o crescimento do animal.
4. Os artrópodes movem-se através de apêndices com juntas articuladas e não pela deformação do corpo (como os anelídeos).
5. O celoma tornou-se vestigial nos adultos e a musculatura é organizada como feixes de músculos: a) músculos extrínsecos: músculos que se ligam nos apódemas do exoesqueleto e se ligam aos apêndices, b) músculos intrínsecos: músculos que ligam artículos dentro do mesmo apêndice.
6. O sistema circulatório é aberto e consta de uma hemocele, com um coração dorsal tubular que pode portar artérias. O coração tem um par de ostíolos (aberturas) laterais por segmento e permitem a passagem do sangue no coração do seio pericárdico adjacente para dentro do coração e ao fechar os ostíolos e contrair o coração o sangue é bombeado para as extremidades do corpo. A hemocianina é o pigmento respiratório mais comum.
7. A excreção é realizada por glândulas localizadas na base das cochas e túbulos de Malpighi nos queliceriformes; glândulas verdes ou antenais nos crustáceos, túbulos de malpighi, nos hexápoda e miriápoda. O escreta mais comum é o ácido úrico.
8. Os órgãos dos sentidos geralmente envolvem alguma especialização do exoesqueleto, os quais permitem o monitoramento dos estímulos ambientais. Pelos ou cerdas são os tipos de sencilas mais comuns nos artrópodes. Apresentam ainda olhos simples (ocelos 1 a 3 ocelos) e olhos compostos.
9. Os artrópodes são dióicos. copula e a fertilização interna são características da maioria das espécies com muitos apêndices envolvidos na corte e na transferência do esperma (espermatóforo).
RESUMO DOS ARTRÓPODES
QUELICERIFORMES
https://youtu.be/QQfnurYNscw
CHELICERATA
Aranhas, escorpiões, opiliões, ácaros
CRUSTACEA
https://youtu.be/-bxRxD4mV1k
Crustacea
Caramarão, lagosta, tatuzinho de jardim
HEXAPODA ou INSECTA
https://youtu.be/bhOP5vZnTmk
HEXAPODA ou INSECTA
Borboletas, besouros, mosca, mosquitos
MYRIAPODA
https://youtu.be/jFq6LPy-0_M
MYRIAPODA
(Chilopoda e Diplopoda)
HOMOLOGIA
EXERCÍCIOS
1) Leia a tirinha de Fernando Gonzales abaixo e explique qual o contexto da tirinha sob o ponto de vista do desenvolvimento dos artrópodes.
Outras tirinhas para vc pensar
2. (Fuvest-SP) Metamorfose é a transformação do estágio jovem para o adulto. Alguns insetos têm metamorfose completa (holometábolos), em outros a metamorfose é incompleta (hemimetábolos). Quais insetos exemplificam o primeiro e o segundo tipo de metamorfose, respectivamente?
a) Gafanhoto e libélula.
b) Borboleta e barata.
c) Mariposa e abelha.
d) Percevejo e mosquito.
e) Besouro e mosca.
3. (UEL-2004) Anelídeos e artrópodes possuem características anatômicas e fisiológicas comuns, o que reforça a hipótese de parentesco evolutivo entre esses grupos de invertebrados. Assinale a alternativa que apresenta, corretamente, duas dessas características comuns:
a) Cordão nervoso dorsal e respiração cutânea.
b) Cordão nervoso ventral e corpo segmentado.
c) Vaso sanguíneo dorsal e respiração traqueal.
d) Vaso sanguíneo ventral e corpo segmentado.
e) Cordão nervoso ventral e vaso sanguíneo ventral.
4. (Unesp) Uma determinada moléstia que pode causar lesões nas mucosas, pele e cartilagens é transmitida por um artrópode e causada por um protozoário flagelado. Os nomes da doença, do artrópode transmissor e do agente causador são, respectivamente:
a) leishmaniose, mosquito anófeles e Leishmania brasiliensis.
b) úlcera de Bauru, mosquito cúlex e Plasmodium vivax.
c) doença do sono, mosca-tsé-tsé e Trypanossoma cruzi.
d) doença de Chagas, barbeiro e Trypanossoma gambiensis.
e) úlcera de Bauru, mosquito flebótomo e Leishmania brasiliensis.
5. (FUVEST)
a) Dê duas características comuns aos crustáceos, insetos e aracnídeos.
b) Dê uma característica própria de cada um desses grupos.
6. (UNESP) Insetos e crustáceos pertencem ao filo Arthropoda. Descreva três características que os diferenciam.
7. (UNESP) - O camarão e a abelha são animais pertencentes ao mesmo filo, embora separados em classes distintas. Cite:
a) duas características que permitam agrupá-los no mesmo filo;
b) duas características que os separam em classes distintas.
8. (FUVEST) Os animais que conversam na tira abaixo pertencem ao filo dos artrópodes. Apesar de carica¬turas, é possível perceber três características externas que os distinguem de qualquer inseto.
a) A que classe pertencem os animais citados?
b) Quais as três características de sua morfologia externa que os distinguem dos insetos?
LEPIDOPTERA
A, B. P. dido dido, Rio Madre de Dios, Perú. C. P. dido dido, Mato Grosso Brazil. D. mating of P. diatonica. E–F. Larva of P. dido chocoensis, Colombia. G–H. Larva of P. pygmalion, Villavicencio, Colombia. I. egg of P. dido chocoensis. J. Pupa of P. dido dido, Villavicencio, Colombia. K. Pupa of P. andrei, Fr. Guiana. L. Larva of P. diatonica, ACG Guanacaste, Costa Rica. M. Passiflora vitifolia, host plant of P. dido chocoensis, Rio Anchicayá, Colombia. Photos: Adrian Hoskins, H. Koloska, Gill Carter, B. Jimenez, Gregory Nielsen, Daniel Janzen, Christian Brevignon, Fredy Montero, Luis M. Constantino.
Os lepidópteros são a segunda ordem mais numerosa dento do subfilo Hexapoda. Pode se dizer que depois dos coleópteros são os animais mais numerosos do planeta. As borboletas e mariposas fazem parte de diversas cadeias tróficas, regulando populações de plantas nativas e servindo de alimento para outros organismos como pássaros e outros hexápodes. No Brasil há uma grande diversidade de espécies de lepidópteros, sendo que muitos são endêmicos.
São insetos de grande importância econômica, pois a maioria das larvas é herbívora (fitófaga) e muitas espécies se tornaram "pragas" de cultivos agrícolas (especialmente mariposas). Contudo, os imagos (adultos) são agentes polinizadores essenciais e fundamentais para o equilíbrio dinâmico dos ecossistemas; são bioindicadores (servem como indicadores da qualidade ambiental), no monitoramento da diversidade biológica de um ecossistema, na integridade de paisagens e uso sustentável de recursos naturais (Brown.
Dryas iulia (Fabr., 1775)
Dryas iulia (Fabricius, 1775) distribui-se do sul dos
Estados Unidos ao norte do Uruguai e Argentina (EMSLEY,
1963; DE VRIES, 1987). Doze subespécies são reconhecidas por EMSLEY (1963), as quais diferem pela coloração
de fundo nas asas, intensidade e extensão das manchas
negras correspondentes e distribuição das androcônias
nas asas anteriores dos machos. Segundo BROWN (1992)
e BROWN & YEPEZ (1984), as populações continentais sulamericanas correspondem predominantemente a Dryas
iulia alcionea (Cramer, 1779).
Esta subespécie é encontrada nos mais variados habitats, predominantemente
naqueles perturbados (perturbação antrópica). Embora suas larvas utilizem diversas passifloráceas como plantas hospedeiras, são
encontradas preponderantemente sobre aquelas do
subgênero Plectostemma Masters (BROWN & MIELKE,
1972; BENSON et al., 1976; PÉRICO & ARAÚJO, 1991; BROWN,
1992).
Dryas iulia alcionea, ovo: 1, vista látero-dorsal; 2, região micropilar; 3, células inferiores; 4, células superiores; 5, aerópila com abertura encoberta; 6, aerópila com tampa decumbente (Ac, abertura da aerópila; Ap, pólo anterior; Hr, carena horizontal; Mp, micrópilas; Pr, peritrema; Ta, tampa da aerópila; Vr, carena vertical; setas indicam posição das aerópilas). Barras: fig. 1, 200 µm; figs. 2, 3, 50 µm; fig. 4, 5 µm; fig. 5, 10 µm; fig. 6, 3 µm.
Bibliografia
Ruppert, E.E. & Barnes, R.D. Invertebrate Zoology. Saunders College Publishing. 1994.
Storer, T. I., & Usinger, R. L. Zoologia Geral. Companhia Editora Nacional. SP. 1978.
Snodgrass, R. E. Principles of insect morphology. Cornell University Press. Ithaca, NY (1935) 1993.
Tiplehorn, C. & Jonnson, N. Estudo dos insetos (tradução da 7ª ed. de Borror & Delong´s introduction to te study of insects). 2011. Cengage Learning. SP
Campbel, N. A. & Reece, J.B. Biologia. Artmed Editora S.A. Porto Alegre.2010
Smart, Paul. The illustrated encyclopedia of the butterfly world. Salamander Books Ltda. London. 1975.
E. S. Chang and D. L. Mykles, Regulation of crustacean molting: A review and our perspectives, Gen. Comp. Endocrinol., 172(3):323–330, 2011 DOI: https://doi.org/10.1016/j.ygcen.2011.04.003
S. P. S. Cheong et al., Evolution of ecdysis and metamorphosis in arthropods: The rise of regulation of juvenile hormone, Integr. Comp. Biol., 55(5):878–890, 2015 DOI: https://doi.org/10.1093/icb/icv066
J. Zhang et al., Functional analysis of insect molting fluid proteins on the protection and regulation of ecdysis, J. Biol. Chem., 289(52):35891–35906, 2014 DOI: https://doi.org/10.1074/jbc.M114.599597
Brown Jr., K. S. & Freitas, A. V. L. 2000. Diversidade de Lepidoptera em Santa Teresa, Espírito Santo. Boletim do Museu de Biologia Mello Leitão, N. Sér. 11/12 pp. 71-118.


























































































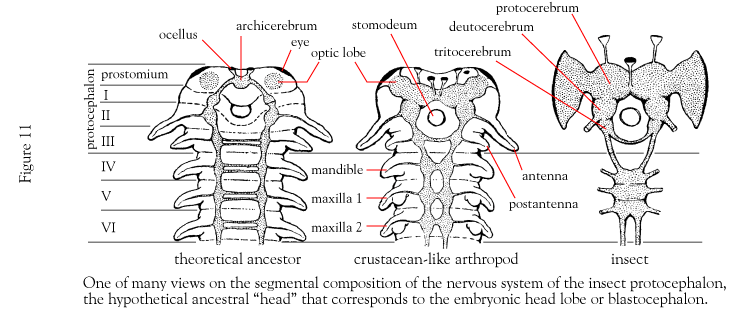
































































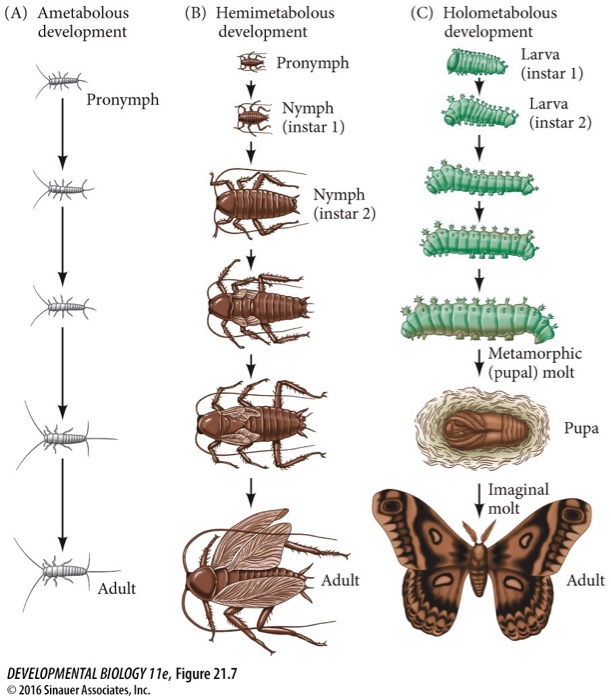





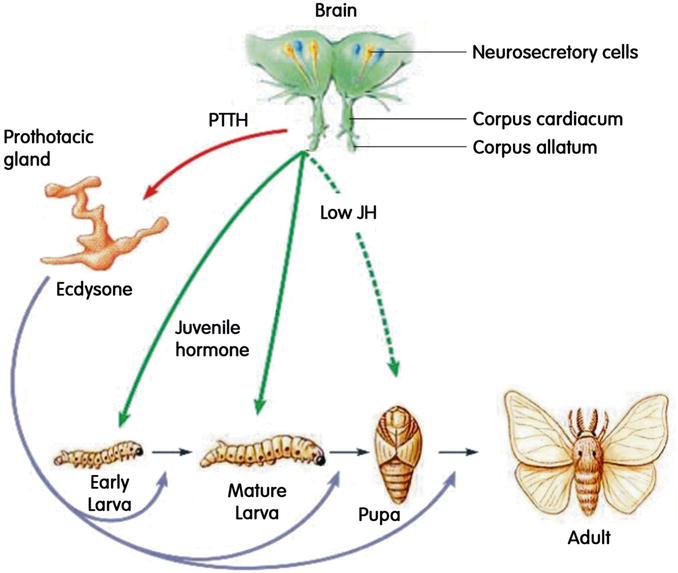























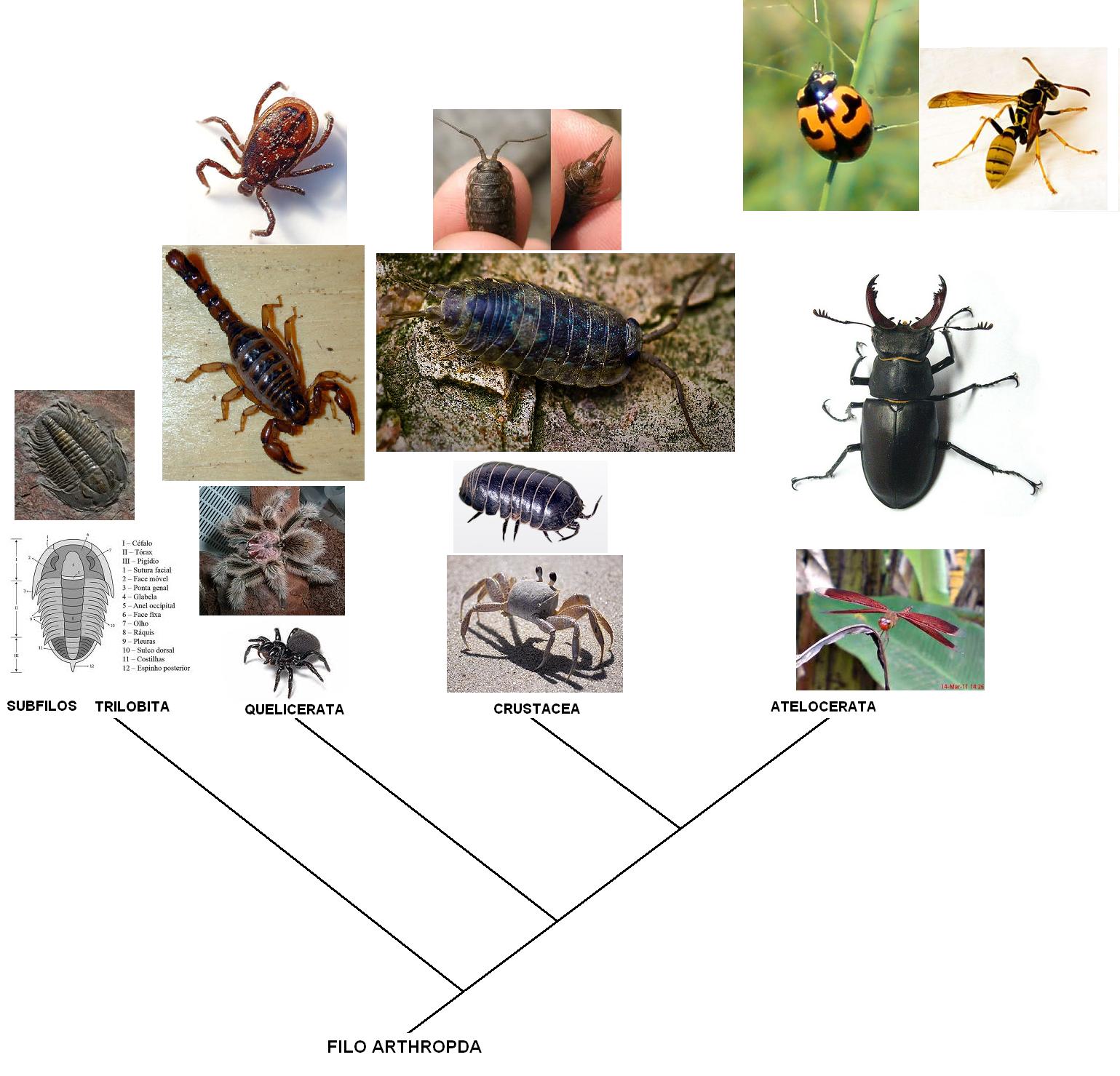









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}