FILO ANNELIDA
ANELÍDEOS
Reino: Metazoa (Animalia)
Sub reino: Eumetazoa
Filo Annelida: do latim annelaatus, anelado, annelus, pequeno anel + o sufixo: idae = semelhante a.
Organismos vermiforme segmentados (metamerizados) corpo alongado e cilíndrico.
O Filo Annelida apresenta mais de 16 mil espécies, são organismos pouco estudados e inclui animais como minhocas, vermes
marinhos e sanguessugas.
O nome do filo como já visto vem do
latim annelatus, annelus, que significa anelzinho mais o sufixo ida (semelhante).
Este filo esta dividido em Polychaeta e Clitellata, e os clitelados abrangem os Oligochaeta e Hirudinea.
Entretanto, essa divisão é puramente didática, uma vez que hoje acredita-se que os Clitellata saíram de dentro dos Polychaetas. Assim, o nome Polychaeta é considerado sinônimo de Annelida. Desta forma, não haveria separação em classes.
Os anelídeos são encontrados principalmente em ambientes marinhos, infaunais, epibentônicos, errantes e sedentários; e ocupam todos os nichos ecológicos.
Podem também serem encontrados de água doce e terrestres. Os Polychaeta são abundantes em "hydrothermal vents" e "cold seeps". As fontes hidrotermais são áreas isoladas onde o fundo do oceano apresenta rachaduras que produzem água aquecida geotermicamente.






Hydrothermal vents ou chaminés hidrotermais
Os organismos que vivem perto dessas aberturas (chaminés hidrotérmicas) são capazes de viver em águas muito quentes (65 °C a 100 °C) que matariam a maioria dos organismos de superfície. Além disso, organismos a essa profundidade são adaptados a pressões extremas. A pressão a cerca de 2.500 metros de profundidade, onde estão localizados os respiradouros, é tão grande que a água superaquecida das nascentes subaquáticas não ferve. De fato, a pressão nessa profundidade impede que a água ferva até atingir cerca de 370 °C.
Em 1977, cientistas em veículos de mar profundo (resistentes a pressões esmagadoras) exploraram fontes termais do fundo do oceano nas Ilhas Galápagos. Neste local foi descoberto uma bactéria que não contém clorofila, mas pode produzir açúcar e compostos orgânicos. Descobriu-se também que a água dos respiradouros é rica em sulfeto de hidrogênio (H2S). A molécula de H2S é rica em energia, mas também é extremamente tóxica para a maioria dos seres vivos. No entanto, as bactérias dessas chaminés são adaptadas para usar a energia dom H2S da mesma maneira que as plantas aproveitam a energia da luz solar. Em vez de realizar a fotossíntese, essas bactérias realizam a quimiossíntese. Essas bactérias servem de alimento para os poliquetos que constituem parte da fauna dessas regiões.
Cold seeps ou infiltrações frias ou escoamentos frios
As emanações frias ou infiltrações frias são outro ambiente do fundo do mar carregado com produtos químicos ricos em energia. Infiltrações frias ocorrem em fissuras ou rachaduras no fundo do mar, causadas pelo movimento das placas tectônicas. O ambiente das fontes hidrotermais e das infiltrações frias é diferente em termos de temperatura e longevidade.
Ao contrário dos produtos químicos ao redor das fontes hidrotermais, as infiltrações frias são semelhantes em temperatura às águas circundantes, ou por vezes até mais quentes, mas com temperaturas muitas vezes mais baixas do que as que ocorrem nas thermal vents.

Exemplo de "cold seeps" e seu entorno
Emanações frias é a designação dada às áreas do leito marinho onde ocorrem emanações, mais ou menos difusas, de sulfureto de hidrogênio (H2S), metano (CH4) e outros fluídos ricos em hidrocarbonetos, frequentemente formando piscinas de salmoura. As emanações frias albergam um bioma que inclui uma grande diversidade de espécies endêmicas. A referência a emanação "fria" não implica que os fluidos emanados sejam mais frios que as águas circundantes, sendo pelo contrário em geral ligeiramente mais quentes, visando apenas distinguir estas emanações das fontes hidrotermais que são sempre muito mais quentes. A presença continuada de emanações frias, por vezes durante períodos geológicos alargados, leva ao desenvolvimento de uma topografia com características únicas, onde as reações entre o metano e a água do mar cria formações e recifes de rochas carbonatadas.
As emanações frias também tendem a ser mais estáveis que as fontes hidrotermais. As fontes hidrotermais são de vida relativamente curta, mas as emanações frias são duradouras. De fato, os vermes que vivem nos ambientes de infiltração a frio estão entre os invertebrados mais longevos do mundo.
A base da cadeia alimentar também é diferente nas infiltrações frias. Nessas infiltrações, o sulfeto de hidrogênio, o metano e outros produtos químicos ricos em hidrocarbonetos lixiviam dos depósitos de petróleo. Esses produtos químicos começaram como matéria orgânica, que foi transformada sob alta temperatura por milhões de anos, eles são uma forma de combustível fóssil. Assim, a energia contida nos produtos químicos de infiltração a frio pode ser rastreada até o sol. Por outro lado, a energia em sulfeto de hidrogênio (H2S) encontrada nas fontes hidrotermais vem do fundo da crosta da Terra.

Esquema de uma "fumarola" ou hydrothermal vent.
Essas fumarolas se formam pela percolação da água que desce pelas
rochas porosas e esquentam próximo a uma fonte de magma.
Ao voltarem a superfície trazem minerais dissolvidos.
Esquema simplificado de uma fonte hidrotermal. A água do mar infiltra-se na crosta oceânica e ao aproximar-se da câmara magmática aquece e é modificada pela troca química com as rochas circundantes. A água do mar alterada, por movimento de convexão, é novamente expelida no fundo oceânico. A pluma crescente mistura-se rapidamente com a água do mar ambiente, baixando a temperatura e a diluição da concentração de partículas. A pluma, menos densa que a água circundante, continua a subir através da água do mar. Uma vez que a densidade da pluma hidrotermal corresponde à densidade da água do mar, a subida pára e começa a dispersar lateralmente. Num cenário como este, 90% dos metais da pluma perdem-se e não formam depósitos de metal. ©Secretariat of the Pacific Community.

Reações químicas que ocorrem próximo a chaminés sulfurosas

Poliquetos marinhos próximos a uma thermal vent
A presença de cerdas é a característica que levamos em consideração para separar os anelídeos em duas classes: Polychaeta e Clitellata.
Os poliquetas (poly = muitas cerdas) são predominantemente marinhos, podem ser infaunais e epibentônicos, mas alguns levam uma vida pelágica. Os oligoquetas e
sanguessugas ocorrem principalmente no solo e em águas continentais. Os
anelídeos podem ter menos que 1 mm de comprimento até mais de 3,0 m.
O menor
anelídeo descrito, um poliqueta, alcança apenas algumas centenas de
micra de comprimento. Também dentro de Polychaeta (Eunicida),
encontramos o maior representante de Annelida: a espécie de Eunice sebastiani que ocorre no litoral norte do estado de São Paulo, e seriamente ameaçada de extinção, alcança mais de 2,0 m de comprimento e 5 cm de diâmetro em sua condição contraída!
Algumas minhocas também podem atingir 1 m de comprimento (Megascolides
australis), e a maior sanguessuga, Haementeria ghilianii sul-americana,
alcança mais de 40 cm de comprimento.

Eunice spp poliqueta marinho
Os anelideos tem grande importancia para a cadeia aliemntar marinha, na formaçãod o humus nos solos agricultáveis
CARACTERÍSTICAS GERAIS
DOS ANELÍDEOS

Corte transversal em um anelídeo oligoqueta.
Plano corporal e origem dos tecidos a partir dos
folhetos embrionários.
Os anelídeos apresentam todos os hábitos alimentares: filtração de suspensões, ingestão de depósitos de material orgânico, detritivoria, herbivoria e carnivoria. são importantes na reciclagem de matéria orgânica (minhocas). São utilizadas na medicina (sanguessugas) para diminuição da pressão sanguínea, regeneração em enxertos e reimplantes de dedos e outros apêndices.
Os poliquetos são extremamente importantes na cadeia alimentar marinha.
As características morfológicas básicas que distinguem os anelídeos são:
animais protostomados, eucelomados e metamerizados (= segmentação não-especializada).
Duas sinapomorfias dentro das classes:
presença de parapódios nos polychaeta e clitelo nos Clitellata.
Os anelídeos compartilham uma origem comum com os moluscos. As evidências são a forma larval trocófora, protostomados com uma cavidade verdadeira (celoma) e a simetria bilateral com a formação de boca e ânus.
Os anelídeos têm um corpo cilíndrico que é dividido em uma série de segmentos separados por um septo de tecido mesodérmico.
O corpo de um anelídeo pode ser dividido em três regiões:
prostômio, tronco e pigídio;
1. Prostômio-Peristômio: boca, cérebro, órgãos dos sentidos;
2. Tronco: maioria dos sistemas, gônadas e apêndices;
3. Pigídio: extremidade posterior do corpo que inclui o ânus.
O prostômio, o peristômio e o pigídio não são segmentos verdadeiros, porque se origem de uma área não segmentar da larva trocófora.
Embora não sejam segmentos verdadeiros eles frequentemente se encontram especializados, nos poliquetas o prostômio geralmente apresenta olhos, antenas, palpos sensoriais, longos tentáculos que servem para captura do alimento. Nos oligoquetas não há nenhuma especialização e o corpo é alongado e cilíndrico, e o prostômio e o peristômio são muito reduzidos. Nos hirudíneos na uma ventosa no anterior circundando a boca que vai até o segmento 5, e uma ventosa posterior que vai do segmento 28 ao 33.

Especializações do prostômio
Todos os segmentos do corpo de um anelídeo são oriundos de uma região da larva trocófora, chamada de zona de crescimento.

Larva trocófora e sua zona de crescimento

Larva trocófora
Charles Robert Darwin trabalhou com esses organismos e chegou a interessantíssimas conclusões, escreveu um livro sobre esses animais onde ele dissecou e desenhou o que viu. Relatou sobre a presença das cerdas (bristles), descreveu a faringe, o esôfago com três pares de glândulas calcíferas e ainda afirmou que não existe nada igual em nenhum outro animal terrestre.
Entre as descobertas de Darwin podemos citar: seu habito noturno; podem rastejar para frente ou para trás por peristalse.
Não possuem olhos mas podem distinguir a noite do dia e respondem rapidamente à qualquer raio de luz; respiram pela pele; são hermafroditas, possuem os dois sexos em um mesmo indivíduo e precisam de um organismo para copular pois não realizam autofecundação; podem viver durante um longo período debaixo d´água; são sensíveis ao ar seco, calor, frio, vibrações no solo e ao serem tocados e manipulados; não possuem dentes na boca e nem garras e são completamente surdos; o seu sentido de cheiro é fraco e limitado à percepção de certos odores; eles são onívoros e preferem alguns tipos de alimento a outro. Eles umedecem os alimentos antes de engolir com um fluido, o qual é da mesma natureza que a secreção digestiva do pâncreas de animais superiores.
Eles são canibais e podem comer minhocas mortas.
Podem viver por um período de tempo de até 16 anos.
Eles escavam as tocas empurrando o solo para os lados e engolindo; eles expelem os dejetos fora de suas tocas. Estes são completamente misturados com secreções intestinais e favorecem o crescimento de plantas, além de serem ricos em carbonato de cálcio; as minhocas são comidos por aves;
Eles regularmente “aram” a terra e misturam o solo da superfície com o solo de camadas mais profundas.
Quem se interessar em ler o incrível livro de Darwin aqui esta o endereço para baixá-lo em pdf gratuitamente.
IMPORTÂNCIA DAS MINHOCAS
NA FERTILIDADE DO SOLO
"It may be doubted whether there are many other animals which have played so important a part in the history of the world." (Charles Darwin, 1881)
As minhocas estão entre os primeiros animais presentes na Terra há 600 milhões de anos e sua importância para a fertilização dos solos já é conhecida há mais de dois mil anos, quando o filósofo Aristóteles as definiu como "arados da terra" por sua capacidade de descompactar os terrenos mais duros.
O naturalista Charles Darwin, autor da Teoria da Evolução das Espécies, dedicou mais de 40 anos aos estudos e pesquisas sobre a vida das minhocas e suas funções na natureza que resultaram no livro "A formação do húmus através das ações das minhocas", publicado em 1881, menos de um ano antes de sua morte, esse livro é bem ilustrado com os desenhos do próprio Darwin e seu filho Francis.
Em algumas espécies, uma única minhoca chega a movimentar de duas a 250 toneladas de hectares de solo. Ao abrir galerias no solo e alcançar seu alimento, que será ingerido, triturado, misturado com carbonato de calcio e enzimas e por fim expelidos em forma de bolotas fecais ou grumos, o húmus de minhoca.
A composição química desse humus contém matéria orgânica (42 a 56%), potássio (1,44 a 2,23%), cálcio (5,44 a 7,26%), magnésio (0,88 a 1,32%), ferro (0,82 a 1,84%), fósforo (1,42 a 3,82%), nitrogênio (1,66 a 2,04), manganês (552 a 767), zinco (418 a 1235) e cobre (193 a 313).




Espécies
Aqui no Brasil, são nativas: a minhoca brava (Pheretyma hawayana), a mansa (Lumbricus terrestris) e o minhocuçu (Rhinodrilus alatus). Entre as espécies exóticas encontramos: minhoca vermelha-da-califórnia ou vermelha-híbrida (Eisenia phoetida), gigante africana (Eudrilus eugeniae) e também a européia (Lumbricus rubelus). Todas essas epécies são usadas nos minhocário para fazer humus e tambem para servir de alimentos a aves, peixes, e rãs.
A formação do solo é um processo de longo prazo determinado em parte pelas condições climáticas e pela natureza do material original (Chesworth, 1992). Envolve a decomposição de minerais primários e a incorporação de matéria orgânica. Darwin (1881) foi um dos primeiros a incluir a biota, especialmente as minhocas, na lista de agentes responsáveis pela formação do solo através do acúmulo de suas fezes e processos de mistura.
O papel potencial das minhocas no desenvolvimento do solo é reconhecido no termo “solos vermiformes” para solos que tenham pelo menos 50% ou mais do horizonte A e> 25% do volume do horizonte B consistindo de estruturas derivadas de minhocas ou animais (tocas, galerias e material fecal). Inicialmente, o termo foi aplicado apenas a Mollisols (solos mollis = moles, típico de campos gramados), mas recentemente foi estendido a outras classes de solo; a relevância desse conceito ainda está sendo discutida, porque a atividade da fauna é observada no perfil da maioria dos tipos de solo (Pop, 1998).
A importância das minhocas no intemperismo químico foi estudada por Darwin (1881) em um experimento em que a cor vermelha do óxido da areia desapareceu após a passagem pelos intestinos das minhocas, provavelmente devido à dissolução do óxido por enzimas acídicas do trato digestivo da minhoca. No entanto, desde o trabalho de Darwin, relativamente pouca pesquisa considerou o papel das minhocas no intemperismo mineral.
Pop (1998) mostrou que as espécies de minhocas de Octodrilus nos Cárpatos Romenos afetam a mineralogia da argila e a formação de ilita no solo (K,H3O)(Al, Mg, Fe)2(Si, Al)4O10[(OH)2,(H2O)] (WP), processo que leva centenas de milhares de anos na ausência de biota.
Em experimentos de laboratório, Carpenter et al. (2007) mostraram que a minhoca epigênica Eisenia veneta Rosa acelerou o tempo de intemperismo da anortita, biotita, esmectita e caulinita; a esmectita foi transformada em ilita e a caulinita reagiu para produzir nova fase mineral (Carpenter et al., 2007).
Sejam as minhocas, os microrganismos estimulados no intestino (Brown, 1995) ou uma ação coletiva de ambos os organismos responsáveis pelo efeito do intemperismo mineral ainda está aberta ao debate.
Em comparação com o intemperismo mineral, o papel da minhoca na formação de húmus foi investigado completamente. O escurecimento do húmus do solo é um processo lento, que envolve reações químicas e atividade microbiana. Esse processo, no entanto, pode ser acelerado por minhocas que preparam misturas de resíduos e litter,(serrapilha) compostas de folhas fragmentadas e maceradas e partículas finas do solo para ataque microbiano. É bem conhecido pelos produtores de vermecompostagem que o húmus pode ser obtido a partir de matéria orgânica dentro de alguns meses (Edwards et al., 2011).
Imaginemos um metro quadrado de solo superficial (em planta) contendo 10 Lumbricus terrestris, cada um com uma vida útil de cinco anos e cada um homogeneizando 1 cm2 de solo durante a vida toda.
Sabe-se que Lumbricus terrestris vive por 12 a 16 anos em condições de laboratório e a área de seção transversal do solo homogeneizado em tocas é visivelmente mais do que a média de 1 cm2 em muitas casos.
Se um novo indivíduo sempre cavar uma toca nova em solo virgem, 10 cm2 do nosso metro quadrado de solo superficial será homogeneizado a cada 5 anos.
Portanto, todo o metro quadrado levaria 5.000 anos para ser homogeneizado. Algum material neolítico e todo o material anterior que não estivesse profundamente enterrado seria potencialmente afetado. Isso ainda é subestimado, pois se aplica apenas à mistura pela população de Lumbricus terrestris. Uma contagem de espécies de minhocas restantes pode muito bem adicionar mais 200 a 300 indivíduos por metro quadrado (ver Russell, 1973: 191; Lee, 1985: 90), alguns dos quais são espécies grandes, como Aporrectodea longa e A. caliginosa, que também penetram profundamente solo e apresentam galerias permanentes (Edwards & Bohlen, 1996).
Em síntese as minhocas desempenham papel fundamental na formação e manutenção da estrutura do solo. Através do processo de bioturbação, (Bioturbação é um processo de alteração físico-química do solo a partir da ação de organismos, como minhocas e térmitas. Esse processo é importante para a manutenção das condições do solo e o fornecimento de água e nutrientes para as plantas) as minhocas formam agregados, alteram a porosidade, aeração e infiltração de água (LAVELLE, 1988). Para sobreviverem num ambiente com pouco alimento e de baixa qualidade, elas desenvolveram uma relação estreita com a microbiota do solo, formando uma simbiose com bactérias no intestino, que permite melhor aproveitamento dos resíduos orgânicos ingeridos (LAVELLE et al., 1995). No intestino há estimulação da atividade microbiana, mineralização de nutrientes, decomposição de MO, reorganização de partículas minerais, e alteração do pH por adição de carbonato de cálcio (CaCO3) (BROWN et al., 2000). Portanto, a passagem do solo pelo trato intestinal das minhocas pode ter um efeito importante sobre a disponibilidade de diversos nutrientes (especialmente o P) para as plantas, e consequentemente sobre a fertilidade do solo (LEE, 1985) (Ferreira, 2020).
Muitas espécies de minhocas, incluindo Pontoscolex corethrurus, possuem ainda a capacidade de sintetizar e secretar carbonato de cálcio (CaCO3) (CANTI; PIEARCE, 2002), além de manterem uma relação simbiótica com fungos micorrizicos (Novais, 2019). A produção de carbonato de cálcio amorfo (CaCO3), ocorre em um órgão especializado das minhocas, as glândulas calcíferas (DARWIN, 1881; CANTI; PIEARCE, 2002). Nessas glândulas é formado um líquido leitoso que compreende micro esférulas amorfas de CaCO3. Acredita-se que essas micro-esférulas se cristalizam para formar grânulos de calcita, aragonita e valerita (GAGO-DUPORT et al., 2008; LEE et al., 2008). Os grânulos de calcita são transferidos das glândulas para o esôfago da minhoca, e a partir daí movem-se para parte final do intestino e são expulsos nos coprólitos (James, 2004; Versteegh et al., 2017, Ferreira, 2020).

Livro de Charles Darwin sobre minhocas
CARACTERÍSTICAS
1) TRIBLÁSTICOS
Ectoderma
Mesoderma
Endoderma
2) SIMETRIA BILATERAL
Só é possível passar um eixo que divide o corpo em metades iguais.
3) PROTOSTÔMIOS
O blastóporo da gástrula dá origem à boca.
Protostomia, grupo de animais, que inclui os platelmintos, nematoides, moluscos, anelídeos, artrópodes. A boca dos Protostômios (proto: primeiro; estoma: boca) se desenvolve da primeira abertura para o intestino embrionário, i.e., do blastóporo.
O celoma (cavidade do corpo) forma-se a partir de uma divisão no mesoderma embrionário. Além disso, o destino celular é fixado na primeira clivagem do embrião inicial, de modo que células separadas não podem formar gêmeos.
As formas larvais, se presentes, são chamadas trocóforos.
A Protostomia constitui uma das duas divisões dos celomados: animais com cavidade corporal ou celoma, a outra divisão constituí os deuterostômios. Por isso os protostômios estão incluídos no clado lofotrocozoa.
4) CELOMADOS, ESQUIZOCELOMADOS
Os anelídeos apresentam como cavidade corporal um celoma verdadeiro do tipo esquizoceloma, i.e., sua origem é de células que migram do endoderma para dentro da blastocele. Essas células se organizam em bolsas dentro da blastocele e crescem ate coalescerem (unem-se formando uma nova cavidade, ou revestindo a antiga blastocele).
O esquizoceloma é então uma cavidade corporal preenchida pelo fluido celômico. Este fluído facilita o transporte interno de substâncias nutritivas e células. Além disso, a presença de um celoma verdadeiro confere vantagens funcionais aos animais que o possui, por ex., a separação entre o sistema digestório e a parede corporal permite que o animal se movimente mais eficientemente, no sentido de que pode se mover sem interferir com os eventos que ocorrem no tubo digestivo.
O surgimento do celoma (esquizoceloma) também permitiu o desenvolvimento de vários órgãos que necessitam de espaço, como as gônadas e órgãos excretores, que se localizam no interior do celoma.
Assim, uma das principais funções do celoma e de seu fluido interno é servir como um esqueleto hidrostático, o qual fornece o apoio para a contração muscular. O que confere maior eficiência, força e precisão aos movimentos executados pelo animal.
O esquizoceloma é então uma cavidade corporal preenchida pelo fluido celômico. Este fluído facilita o transporte interno de substâncias nutritivas e células. Além disso, a presença de um celoma verdadeiro confere vantagens funcionais aos animais que o possui, por ex., a separação entre o sistema digestório e a parede corporal permite que o animal se movimente mais eficientemente, no sentido de que pode se mover sem interferir com os eventos que ocorrem no tubo digestivo.
O surgimento do celoma (esquizoceloma) também permitiu o desenvolvimento de vários órgãos que necessitam de espaço, como as gônadas e órgãos excretores, que se localizam no interior do celoma.
Assim, uma das principais funções do celoma e de seu fluido interno é servir como um esqueleto hidrostático, o qual fornece o apoio para a contração muscular. O que confere maior eficiência, força e precisão aos movimentos executados pelo animal.
5) ESTÃO COLOCADOS NO CLADO DOS LOFOTROCOZOA
Os animais protostômios dividem-se em Lofotrocozoa e Ecdisozoa. O clado Lophotrochozoa inclui os Platyhelminthes, Lophophorata, Mollusca e Annelida. Isto porque apresentam larva trocófora e tentáculos ao redor da boca.


Divisão simplificada dos animais triblásticos em protostômios e deuterostômios.
Os protostômios estão divididos em Lofotrocozoa e Ecdisozoa
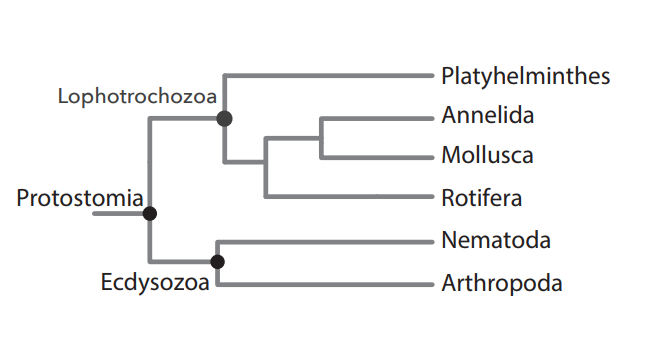
O clado Lofotrocozoa inclui os platelmintos os anelídeos, os moluscos e os rotíferos (não estudados)
Os anelídeos são os primeiros a apresentar metameria, que na zoologia é sinônimo de segmentação (que esta presente em anelídeos, artrópodes e cordata), ou seja, o corpo é formado por repetição, iguais ou semelhantes, de unidades funcionais, os metâmeros, que se repetem ao longo do eixo ântero-posterior do corpo do animal.
Os animais que não desenvolvem essa segmentação corporal são chamados de ametaméricos (como os filos estudados anteriormente).
Os animais segmentados têm tipicamente uma região anterior chamada de prostômio, (ou ácron), um tronco e uma região posterior chamada pigídio ou télson.
O corpo de um anelídeo é caracterizado pela segmentação, na qual a maioria das partes internas e externas é repetida em cada segmento, o que seria uma homologia seriada, i.e., estruturas corporais com as mesmas origens genéticas e de desenvolvimento aparecem repetidamente durante a ontogenia de um organismo, o que resulta em metameria. Os animais que não desenvolvem essa segmentação corporal são chamados de ametaméricos (como os filos estudados anteriormente).
Os animais segmentados têm tipicamente uma região anterior chamada de prostômio, (ou ácron), um tronco e uma região posterior chamada pigídio ou télson.
Essa segmentação do corpo se origina de um desenvolvimento teloblástico, proliferação de bandas mesodérmicas segmentares pareadas a partir de células teloblásticas que ficam na zona posterior de crescimento do embrião (teloblastos são células ligadas ao crescimento, e o crescimento teloblástico resulta na adição de novos segmentos).
Uma das vantagens da metameria está relacionada à locomoção dos animais, visto que o volume de líquido celomático se mantem constante em cada compartimento (metâmero), funcionando como força antagônica contra a qual os músculos (circulares e longitudinais) podem atuar, e, também, ao fato de haver a possibilidade de controlar independentemente cada segmento. Além disso, a segmentação levou a uma repetição de estruturas, o que pode diminuir o impacto de uma danificação corporal, pois os segmentos não danificados podem manter as suas funções praticamente normais e, assim, aumentar a probabilidade do animal superar os danos sofridos e sobreviver.
Relembrando
A metameria verdadeira é aquela em que a segmentação do corpo se desenvolve pela segmentação do mesoderma, ocorre nos anelídeos, artrópodes e maioria dos cordados. Os segmentos são interdependentes, integrados em uma única unidade funcional, e todos trabalham em cooperação, além de que novos segmentos são formados na extremidade posterior do corpo (em frente ao segmento anal).
A metameria verdadeira é aquela em que a segmentação do corpo se desenvolve pela segmentação do mesoderma, ocorre nos anelídeos, artrópodes e maioria dos cordados. Os segmentos são interdependentes, integrados em uma única unidade funcional, e todos trabalham em cooperação, além de que novos segmentos são formados na extremidade posterior do corpo (em frente ao segmento anal).
Já o "pseudometamerismo" diz respeito à segmentação do corpo quando ocorre pela segmentação do ectoderma, e é vista em vermes (Platelmintos). Nesses, os segmentos são funcionalmente independentes e não há cooperação entre eles, e os novos segmentos são formados na parte anterior do corpo atrás do escolex, como exemplo temos a Taenia spp.
Uma das vantagens da metameria está relacionada à locomoção dos animais, visto que o volume de líquido celomático se mantem constante em cada compartimento (metâmero), funcionando como força antagônica contra a qual os músculos (circulares e longitudinais) podem atuar, e, também, ao fato de haver a possibilidade de controlar independentemente cada segmento. Além disso, a segmentação levou a uma repetição de estruturas, o que pode diminuir o impacto de uma danificação corporal, pois os segmentos não danificados podem manter as suas funções praticamente normais e, assim, aumentar a probabilidade do animal superar os danos sofridos e sobreviver.
Outra vantagem seria a modificação de certas porções do corpo para o desempenho de funções especializadas, o que está relacionada a três processos: a restrição de certas estruturas segmentares a apenas alguns segmentos (por ex.: as gônadas dos Oligoquetas restringem-se a apenas alguns segmentos, os segmentos genitais), a divergência, que é a existência de estruturas segmentares que estão adaptadas ao desempenho de várias funções (por ex.: apêndices adaptados para a alimentação, trocas gasosas, reprodução, etc.); e a fusão de segmentos dando origem a zonas diferenciadas.
A segmentação ou metameria é característico de animais cujos corpos são divididos em séries longitudinais de unidades repetidas ou segmentos (metâmeros ou segmentos).
Uma das vantagens mais importante da segmentação é que esta divide o corpo em uma série de compartimentos, cada um dos quais pode ser regulado mais ou menos independentemente. Assim, a evolução da segmentação, como a evolução da multicelularidade e da colonialidade proveu uma rede de especialização.
Em animais coloniais (cnidários por ex.), tal especialização é aparente no polimorfismo dos zoóides, enquanto nos animais segmentados ela resulta em uma especialização regional dos segmentos do corpo.
O mais simples exemplo de especialização regional é encontrado em anelídeos nos quais o corpo pode ser fracamente dividido em cabeça, tórax e abdomen. Essas mesmas divisões são mais proeminentes nos ARTHTROPODA (como insetos e muitos crustáceos). No caso extremo, por exemplo, nos vertebrados superiores, mesmo o arranjo segmental dos músculos é obscurecido pela especialização regional (do corpo).
A especialização regional do corpo é o resultado de três processos:
1- restrição
2- divergência e
3- fusão.
1. Restrição
O mais simples exemplo de especialização regional é encontrado em anelídeos nos quais o corpo pode ser fracamente dividido em cabeça, tórax e abdomen. Essas mesmas divisões são mais proeminentes nos ARTHTROPODA (como insetos e muitos crustáceos). No caso extremo, por exemplo, nos vertebrados superiores, mesmo o arranjo segmental dos músculos é obscurecido pela especialização regional (do corpo).
A especialização regional do corpo é o resultado de três processos:
1- restrição
2- divergência e
3- fusão.
1. Restrição
Certas estruturas segmentais tornam-se restritos à apenas alguns segmentos, por ex.: as gônadas são geralmente restritas à uns poucos segmentos genitais especializados.
2. Divergência
Outras estruturas segmentares podem ser retidas em todos segmentos mas podem estruturalmente divergir dependendo do local em que se encontram, e adotar funções diferentes. Tal divergência é comum entre apêndices segmentados alguns dos quais podem ser especializados para locomoção, enquanto outros funcionam, por ex.: para raspar, mascar, realizar trocas gasosas.
3. Fusão
A especialização também resulta da fusão dos segmentos. Embora a fusão possa ocorrer em qualquer lugar ao longo do comprimento do animal segmentado é comumente expresso como fusão anterior de um ou mais segmentos com um prostômio para formar uma cabeça complexa.
A cabeça de um poliqueta como o Nereis sp. é formada de um prostômio e no mínimo dois segmentos enquanto que a cabeça de uma mosca das frutas (Drosophila melanogaster) é composta de cinco segmentos.
A cabeça de um poliqueta como o Nereis sp. é formada de um prostômio e no mínimo dois segmentos enquanto que a cabeça de uma mosca das frutas (Drosophila melanogaster) é composta de cinco segmentos.
7) PAREDE DO CORPO
Iniciando de fora para dentro teremos:
Cutícula
Epiderme glandular
Camada muscular circular
Camada muscular longitudinal
Peritônio (mesoderma) que forra todo o grande celoma
Na linha mediana dorsal o peritôneo se abre para segurar o vaso sanguíneo dorsal e se fecha novamente e logo abaixo do vaso ele se abre novamente para permitir a passagem do tubo digestório; abaixo deste ele se abre novamente para deixar passar o cordão nervoso ventral, os dois nefróstomas e o cordão nervoso ventral. Alem disso o celoma esta dividido em duas metades a metade esquerda e a direita são independentes.

Parede do corpo de um anelídeo

Parede do corpo e septo formado pelo peritônio (mesoderma).
O peritônio que reveste o tubo digestório se chama peritônio visceral,
e o peritônio que reveste a parede do corpo é chamado de peritônio parietal.
Corte transversal de uma minhoca
O peritônio que envolve o tubo digestivo é chamado de peritônio visceral e o peritônio que envolve o celoma é chamado de peritônio parietal.
8) MOVIMENTO
A compartimentalização do celoma aumenta a precisão dos movimentos pois pode-se aplicar pressão sobre diferentes zonas do líquido. Ondas de contração muscular, peristálticas, controladas pelo sistema nervoso, passam pelo corpo, alongando e contraindo alternadamente grupos de segmentos, forçando o corpo do animal a deslocar-se, com o auxilio das cerdas que estão ancoradas nas paredes da galeria ou seja, elas dão tração ao animal.
Peristalse: Conjunto de contrações musculares em órgãos ocos ou paredes musculares que delimitam espaços com líquido (segmento, metâmero), cavidade celômica que atua como esqueleto hidrostático, que através de ação sobre o líquido propele o corpo para frente.
Sucessão de ondas de contração muscular involuntária de vários órgãos ocos, ou tubos corporais, especialmente do trato alimentar, onde efetua o transporte de alimentos e resíduos.
O controle do peristaltismo tanto do sistema digestório quanto dos metâmeros dos anelídeos está a cargo do sistema nervoso (gânglios de cada segmento ou do plexo submucoso e plexo mioentérico no sistema digestório).
Do grego antigo, δύναμις περισταλτική, dynamis peristaltikḗ, ação do intestino pela qual seu conteúdo é propelido, de περιστέλλω, peristéllō, peristellein, envolver, de περι- peri-, ao redor + στέλλω, stéllō, stellein, preparar.
As quatro fileiras de cerdas ventrais dos oligoquetas auxiliam no movimento para frente. Estas cerdas quitinosas impedem o animal de deslizar para trás, reforçando o movimento das camadas musculares circulares e longitudinais.
Cerdas importantes no movimento e órgãos internos de oligoqueta

Esquema do movimento por peristalse em minhocas
A minhoca é um organismo sem membros com um esqueleto hidrostático que se move por peristaltismo. Seu esqueleto hidrostático consiste em uma cavidade corporal cheia de líquido, cercada por uma parede corporal muscular extensível. O animal se move contraindo radialmente a porção anterior de seu corpo, resultando em um aumento no comprimento por meio da pressão hidrostática. Essa região restrita se propaga posteriormente ao longo do corpo do animal. Como resultado, cada segmento é estendido para a frente, então relaxa e entra em contato novamente com o substrato. As cerdas em forma ancoram cada segmento ao substrato, impedindo o deslizamento para trás. A contração e a distensão de músculos circulares e longitudinais sobre o esqueleto hidrostático e o auxilio das cerdas protraídas e "ancoradas" nas paredes da galeria, faz com que o movimente para frente ou para trás seja facilitado.

Movimento em poliqueta marinho

Movimento serpenteante em poliqueta
9) CERDAS OU QUÊTAS
Estruturas quitinosas que tem função na locomoção e ancoragem nos túneis (minhocas terrestres) e auxiliam no deslocamento e na respiração branquial nos poliquetas (marinhos) que apresentam cerdas modificadas chamadas de parapódios. São úteis para diferenciar os grupos de anelídeos.

Cerdas em oligoquetos usadas para ancorar o corpo contra as paredes do túnel.
Forma epítoca de Nereis succinea mostrando as cerdas bem desenvolvidas

Cerdas e órgãos internos em poliqueta marinho

Parapódios
10) FORMA DO CORPO
Apresentam corpo alongado e cilíndrico, divido em anéis ou metâmeros, estes separados por septos.


Metameria
11) SISTEMA CIRCULATÓRIO
Sistema circulatório fechado, formado por vasos sanguíneos interligados, e vasos segmentares. O vaso dorsal leva o sangue em direção anterior e é contrátil, ele pulsa. E o vaso ventral leva o sangue da região anterior para a posterior. Em cada segmento do corpo há vasos capilares que formam microcircuitos, do vaso dorsal sai uma rede de vasos capilares que envolvem o intestino absorvem nutrientes e chegam ao vaso ventral. Do vaso ventral sai uma rede de capilares que chamada de plexo epidérmico, que vai até a superfície da pele faz as trocas gasosas e chega ao vaso dorsal.
Como vimos, o surgimento do celoma ocasionou o aumento do volume e, conseqüentemente, da complexidade corporal. Portanto, nos anelídeos, passou a ser necessário um sistema de transporte mais eficiente que pudesse unir as regiões de obtenção e demanda de nutrientes, levando com rapidez as substâncias essenciais para o metabolismo e retirando aquelas que restaram após o processo metabólico (os excretas).
Em geral, os anelídeos têm um sistema circulatório fechado, que consiste em dois vasos sangüíneos principais (um dorsal e outro ventral) ligados por uma rede de capilares. O vaso sangüíneo dorsal conduz o sangue para a região anterior, enquanto o vaso ventral leva o sangue para a região posterior (Figura 14.6).
O fluxo unidirecional do sangue é garantido pela presença de válvulas nesses vasos sangüíneos. O fluxo do sangue é mantido pela contração dos próprios vasos, principalmente o dorsal. Isso ocorre na maioria dos anelídeos.
Entre as sanguessugas (classe Hirudinea), esse tipo de sistema circulatório fechado está muito reduzido ou mesmo não existe. Nesses animais, o fluido celômico é responsável pela circulação, seja integralmente ou parcialmente.
O fluido alcança os tecidos internos pelos canais e seios celômicos, presentes nos hirudíneos.
Outra característica importante do sistema circulatório dos anelídeos é a presença de pigmento respiratório no sangue da maioria dos poliquetas, oligoquetas, hirudíneos. A hemoglobina é o pigmento mais comum, mas também ocorrem dois outros pigmentos, a clorocruonina e a hemeritrina.
Esses pigmentos podem ocorrer simultaneamente no sangue de um anelídeo.
Há mais de um tipo de hemoglobina, mas em todos há uma ligação das moléculas de oxigênio com os átomos de ferro presentes na molécula desse pigmento. Como a hemoglobina, a clorocruonina (vermelho claro quando contem oxigênio e verde quando esta sem oxigênio) também contém ferro em sua molécula, mas, apesar de ser quimicamente semelhante à hemoglobina, este pigmento possui uma coloração esverdeada (quando sem oxigênio) e encontra-se dissolvido no sangue. A hemeritrina ocorre, no mínimo, em uma espécie de poliqueta. Este pigmento é estruturalmente diferente dos outros dois e se encontra dentro de células, e não em solução no sangue.

Sistema circulatório (vaso dorsal, vaso ventral e micro circuitos de capilares)

12) SISTEMA EXCRETOR
Os anelídeos são animais ureotélicos, i.e., excretam uréia, que é uma substancia solúvel em água, todavia menos tóxica que a amônia.

Protonefrídio em platelmintos e metanefrídio em anelídeos



Metanefrídio de anelídeos
13) SISTEMA NERVOSO E ÓRGÃOS DOS SENTIDOS
O sistema nervoso é formado por um par de gânglios cerebrais suprafaríngeo (ou supraesofágico dependendo da posição) semelhantes a um cérebro que se localiza acima e a frente da faringe (anterior e dorsal). Do cérebro parte um anel circum-faringiano ou circum-esofágico de cordões nervosos ao redor da faringe (ou esôfago), que conecta-se a um primeiro gânglio subfaríngeo ou subesofágico, este conecta a gânglios ventrais em cada segmento. O sistema nervoso é completamente metamerizado. Um único cordão mediano ventral ganglionar com um gânglio para a metâmero inerva todo o corpo do animal.


Gânglios cerebrais ligados por um anel nervoso circum-faringeano que ligam o "cérebro" ao primeiro gânglio sub-faríngeo ou sub-esofágico.

Cordão nervoso mediano ventral com com um gânglio em cada metâmero.

Ocorrem olhos e cerdas sensoriais
14) REPRODUÇÃO
A grande maioria dos poliquetas possui sexos separados (isto é, são dióicos) e liberam seus gametas na água. Após a fertilização externa, forma-se uma larva do tipo trocófora, que permanecerá na coluna d’água por um tempo variado (semanas a meses) até sofrer a metamorfose e se transformar em um juvenil.
O tempo da fase larval varia bastante dentro da classe. Muitas espécies são capazes de se reproduzir assexuadamente, através da fragmentação ou divisão do corpo, em um processo chamado epitoquia.
As minhocas e sanguessugas (classe Clitellata) são hermafroditas e realizam a fertilização cruzada entre dois indivíduos. Não há liberação de gametas no ambiente, portanto, a fecundação dos ovos (ricos em vitelo) ocorre dentro do corpo do animal.
Não há gônadas individualizadas nos poliquetas, os gametas são formados ao longo do peritônio e são liberados pelos nefróstomas. Nos clitelados há gônadas onde se desenvolvem e amadurecem os gametas.

Cópula entre dois oligoqueta (hermafroditas)
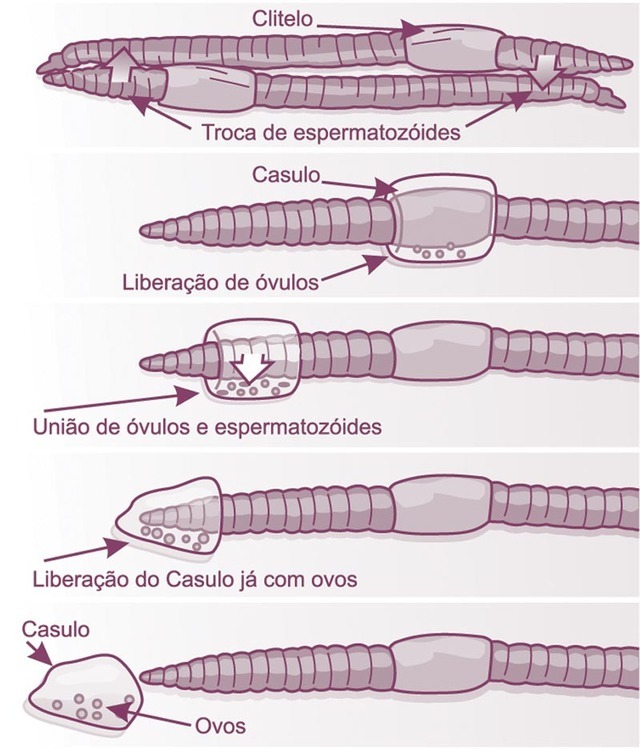
Reprodução nos oligoquetas

Cópula e produção dos casulos
15) SISTEMA DIGESTÓRIO
Existem espécies predadoras, como, por exemplo, os poliquetos carnívoros que possuem mandíbulas para a captura de alimento; outras espécies são comedoras de depósito (comedores seletivos ou não seletivos de depósito), detritívoras, ingerem sedimentos que constitui o solo, as minhocas, que são comedores não seletivo de depósito, possuem também cecos intestinais e tiflossole modificações que aumentam muito a absorção do alimento, por aumentar a área de contato do sedimento com o tecido de absorção; outras espécies se alimentam por um tipo de filtração e assim são suspensívoros.
E ainda temos as sanguessugas que alimentam-se de sangue de outros animais, por sucção.
Possuem também dentes em formas variadas formando mandíbulas; Alguns grupos apresentam glândulas de veneno (Gliceridae), glândulas esofágicas (Nereidae e outros) e glândulas produtoras de muco (vários grupos); A região média do sistema digestivo (estômago) contém células secretoras que produzem enzimas digestivas, enquanto a região posterior (intestino) é ciliada e possui ação peristáltica que facilita a digestão. Entre os poliquetos existe uma grande diversidade de formas alimentares que permitiu explorar quase todas as fontes marinhas: Raptoriais (Filodocidae, Silidae e Nereidae): caçam e detectam as presas por meios químicos ou mecânicos, e evertem a faringe para sua captura; Predadores oportunistas (Polynoidae) esperam as presas em tocas ou tubos; Comedores de depósitos (Arenicolidae, Opheliidae, Maldanidae) ingerem o substrato e digerem o material orgânico nele contido; Suspensívoros (Serpulidae e Sabelidae) utilizam uma coroa de tentáculos peristomiais ramificados (radíolos) para produzir uma corrente de água com alimentos. Parasitas: (Oenonidae) vive no celoma de outros poliquetas; Ichthyotomidae são endoparasitas de enguias marinhas. (Fonte)

Corte longitudinal e transversal e uma minhoca (clitelado)
mostrando os órgão internos

Sistema nervoso, sistema circulatório e sistema excretor.

Sistema digestório de uma minhoca

Sistema digestório e nervoso


Main interactions in the Riftia symbiosis. "HOST" refers to processes in Riftia host tissues, while "SYMBIONT" refers to processes in the bacterial endosymbiont.
A plus sign indicates presumably stimulating interactions, and a minus sign indicates presumably inhibiting interactions. For example, host efforts that protect the symbiont population from oxidative stress, i.e., ROS detoxification and fermentative metabolism (on the right), can promote symbiont biomass production (+). In contrast, host immune system-related proteins and antimicrobial peptides (AMPs) may inhibit symbiont biomass production (−). Circles, where present, indicate that the respective proteins are more abundant in S-rich (energy-rich) specimens (light circles) or S-depleted (energy-limited) specimens (dark circles). The dashed arrow indicates putative transfer of small organic compounds "Milking".
Principais interações na simbiose de Riftia. "HOST" refere-se a processos nos tecidos hospedeiros de Riftia, enquanto "SYMBIONT" refere-se a processos no endossimbionte bacteriano. Um sinal de mais indica interações presumivelmente estimulantes e um sinal de menos indica interações presumivelmente inibidoras. Por exemplo, os esforços do hospedeiro que protegem a população de simbiontes do estresse oxidativo, ou seja, desintoxicação de ROS e metabolismo fermentativo (à direita), podem promover a produção de biomassa de simbiontes (+).
Por outro lado, proteínas relacionadas ao sistema imunológico do hospedeiro e peptídeos antimicrobianos (AMPs) podem inibir a produção de biomassa de simbionte (-). Os círculos, quando presentes, indicam que as respectivas proteínas são mais abundantes em espécimes ricos em S (ricos em energia) (círculos claros) ou espécimes esgotados em S (limitados em energia) (círculos escuros). A seta tracejada indica transferência putativa de pequenos compostos orgânicos "Ordenha".
https://youtu.be/VLCmnLLbj5g
"Verme" Bobbit Poliqueta predador
https://youtu.be/7PMmrrmG1uc
Bobbit o poliqueta predador
16) TROCAS GASOSAS
As trocas gasosas ocorre nos anelídeos através da parede corporal, que é, em geral, bastante permeável. Entretanto, devido ao aumento da complexidade corporal, como vimos anteriormente, a troca de gases respiratórios por simples difusão pela parede do corpo não é suficiente para manter o seu funcionamento. Em muitas espécies, especialmente de poliquetas, as trocas gasosas ocorre em expansões corporais especializadas, que possuem grande vascularização de capilares sangüíneos, como é o caso dos parapódios.
Os parapódios funcionam como brânquias e estão presentes em grande número ao longo do corpo do animal. Nas espécies de poliquetas que vivem em tubos (poliquetas sedentários), há o desenvolvimento de tentáculos na região anterior que possuem a função de brânquias, além de realizar a coleta de alimentos.
Os oligoquetas e hirudíneos dependem da difusão de gases pela parede corporal. Por isso, as espécies terrestres estão limitadas aos ambientes úmidos e bem oxigenados, pois necessitam manter a parede corporal umedecida para facilitar a absorção de oxigênio. A presença de vasos sangüíneos próximos da parede do corpo e de pigmentos respiratórios no sangue aumentam a eficiência da troca gasosa e do transporte do oxigênio para os tecidos mais internos.
Bibliografia
(Visitado em 08/VII/2020)
SISTEMÁTICA DO FILO ANNELIDA

POLYCHAETAS
Vermes principalmente marinhos segmentados.
Hábitat embaixo de pedras, buracos temporários ou em tubos permanentes secretados pelo próprio animal.
Os sexos separados, fecundação externa (óvulos e espermatozóides na água do mar).
Desenvolvimento indireto com uma larva trocófora livre-natante;
Prostômio (porção anterior) com apêndices sensitivos (olhos, palpos e tentáculos) e cérebro;
Peristômio logo após o prostômio (é o 1° segmento verdadeiro), porta a boca no lado ventral;
Pigídio (parte terminal do corpo) onde encontra-se o ânus;
Corpo metamerizado, com muitos metâmeros, metâmeros com numerosas cerdas implantadas em um par de parapódios laterais.
O próprio nome diz Polychaeta: apresentam muitas cerdas. Todos são marinhos ex.:
Nereis sp
Eunice sebastiani, espécie endêmica do litoral de São Paulo.
A boca localiza-se ventralmente entre o prostômio e o peristômio. Enquanto o pigídio é um segmento terminal, onde fica o ânus. Os poliquetas distribuem-se na coluna de água desde entre-marés até grandes profundidades (mais de 4 mil metros). Todavia ha registros de poliquetas nas fossas Marianas ha mais de 10.000 m de profundidade.
De fora para dentro teremos a cutícula iridescente, a epiderme é um epitélio colunar muitas vezes ciliado em certas partes do corpo; logo abaixo desta, localiza-se uma camada de tecido conjuntivo, musculatura circular e grossos músculos longitudinais. Os músculos circulares não são contínuos, sendo interrompidos na altura dos parapódios. O revestimento interno é o peritônio, que circunda espaços celômicos e reveste as superfícies dos órgãos internos.
O celoma está arranjado segmentarmente em direito e esquerdo. Mesentérios dorsal e ventral separam os membros de cada par de celomas, e septos musculares intersegmentares isolam cada par, ao longo do corpo. Os septos intersegmentares de alguns animais foram secundariamente perdidos ou estão perfurados, de modo que o fluido celômico fica contínuo entre os segmentos. Em muitos poliquetas pequenos, o revestimento celômico é perdido.
Além dos músculos já mencionados, outros servem para retrair partes protaíveis e eversíveis, por exemplo, brânquias e faringe, e para manobrar os parapódios.
Os parapódios são evaginação da parede do corpo que se movem primariamente por músculos diagonais. Os parapódios grandes contêm um par de hastes de sustentação quitinosas e escleroproteínicas chamadas acículas, nas quais alguns músculos se inserem. As cerdas também são supridas por músculos e podem geralmente ser retraídas e estendidas

Detalhe de um parapódio


Prostômio, peristômio e pigídio

Corte transversal mostrando músculos diagonais
Em geral, poliquetas são animais cavadores que apresentam uma musculatura circular bem
desenvolvida e septos incompletos, onde o fluido celomático (que funciona
como esqueleto hidrostático) é compartilhado pelos segmento, uma vez que possuem menos septos.
O deslocamento se dá por movimentos serpenteantes que é característico dos poliquetas e também por peristalse.
As trocas gasosas são realizadas por meio de cirros presentes nos parapódios ou associados a brânquias, sendo que estas aparecem pela primeira vez na história evolutiva. Os poliquetas são divididos em errantes e sedentários.
Os errantes são epibentônicos que nadam ativamente no substrato marinho a procura de alimento, podem ser predadores, carnívoros, herbívoros, detritívoros, ou onívoros. Os sedentários constroem tubos ou ficam enterrados no sedimento sendo portanto comedores de depósitos seletivos ou não seletivos ou suspensívoros.

Poliqueta suspensívoros
Um sabelídeo, poliqueto suspensívoro, expande sua coroa de radíolos para se alimentar. Cílios dos radíolos e das pínulas promovem circulação de água por entre eles; as partículas de alimento são aglutinadas em muco e conduzidas em fendas ciliadas até a boca. As partículas de areia descartadas pelos radíolos são armazenadas em um saco ventral e utilizadas na construção do tubo. B) Detalhe de um radíolo, mostrando os tratos ciliares e o mecanismo de seleção de partículas por tamanho. (Fonte: modificado de Hickman et al., 2004; elaborado por USP/Univesp).

Poliqueta comedores de depósito não seletivo
Arenicola escava uma galeria em forma de “L” na areia ou lodo e promove circulação de água no interior dela a partir de movimentos peristálticos. A água entra pela abertura da galeria e é forçada para seu fundo cego, sendo filtrada na areia dessa região. Essa areia, contendo grande quantidade de material filtrado e de sedimento, é, então, ingerida pelo poliqueto. (Fonte: modificado de Hickman et al., 2004; elaborado por USP/Univesp).


Chaetopterus cf. appendiculatus: (A) poliqueta completo; (B) tubo; (C) detalhe da superficie do interior do tubo. Chaetopterus sp.: (D) animal inteiro; (E) tubo; (F) detalhe da superfície interna do tubo.

Chaetopterus sp. vive em um tubo em forma de “U” aberto em ambas as extremidades. Em determinados segmentos, os notopódios estão modificados como leques que criam a corrente de água que flui através da galeria. O 12° segmento possui parapódios que secretam uma bolsa de muco; a água que flui através dessa bolsa mucosa é filtrada. Periodicamente, a bolsa repleta de partículas filtradas é ingerida pelo poliqueta. (Fonte: modificado de Hickman et al., 2004; elaborado por USP/Univesp).

Amphitrite, um poliqueto tubícola cujos longos tentáculos são estendidos sobre o substrato para captura de alimento. Os cílios e muco dos tentáculos capturam as partículas do sedimento e as conduzem à boca. (Fonte: modificado de Hickman et al., 2004; elaborado por USP/Univesp). A maioria dos poliquetos tubícolas e cavadores é suspensívora ou comedora de depósito, utilizando o mecanismo básico ciliar e mucoso para obtenção de alimento. Por exemplo, em alguns poliquetos tubícolas, como Amphitrite, há longos tentáculos (apêndices modificados do protostômio) que são estendidos sobre o substrato (lodo ou areia); cílios e muco dos tentáculos capturam as partículas do sedimento e conduzem-nas à boca. (Fonte: modificado de Hickman et al., 2004; elaborado por USP/Univesp).

Terebella lapidaria (Linnaeus, 1767)
Comedores de depósito seletivo
Quanto a reprodução podem se reproduzir por fragmentação seguido de regeneração, ou ocorre uma divisão a cada poucos segmentos onde ocorre a formação de uma região anterior e uma região posterior, e normalmente ficam unidos formando uma fila de indivíduos, todos geneticamente iguais. Na reprodução sexuada os gametas são produzidos pelo peritôneo (mesoderma que reveste o celoma) e maturam no próprio celoma, sendo eliminados pelos nefridióporos ou pela ruptura da parede do corpo, o que leva a morte do indivíduo parental. Na fecundação ha formação de uma larva trocófora como nos moluscos (os anelídeos são o grupo irmão dos moluscos; trocozoa).

grupos irmãos anelídeos e moluscos
Há ainda a epitoquia e enxameamento. Em diversos poliquetas marinhos é comum a ocorrência do fenômeno reprodutivo de epitoquia, onde um indivíduo não reprodutor epibentônico, que se alimenta normalmente no substrato marinho, sem adaptações para nadar na coluna d'água, chamado de átoco, dá origem a um indivíduo reprodutor pelágico denominado epítoco. O epítoco tem cerdas apropriadas para natação e existe única e exclusivamente para a reprodução.
A epitoquia se assemelha a uma metamorfose e envolve mudanças no animal para a natação e reconhecimento do parceiro sexual, além da produção de gametas e maturação sexual, ou o animal dá origem ao epítoco através de brotamento. Este apresenta olhos maiores, pois necessita nadar na coluna d'água.
Os epítocos então vão para a coluna d'água e nadam para a superfície simultaneamente onde ocorre a cópula e assim liberam seus gametas (óvulos e espermatozoides). Este comportamento sincronizado é conhecido como enxameamento e tem grande importância na garantia da variabilidade genética e no sucesso da fecundação. Há um componente relativo a periodicidade lunar, pois esse fenômeno acontece normalmente ha milhões de anos na lua cheia da primavera. (A produção dos epítocos é regulada pela lua, e o enxameamento ocorre na primeira lua cheia da primavera). Na polinésia ocorre um festival onde as pessoas presenciam o fenômeno e depois capturam os epítocos para comer, que é considerado uma iguaria.
O estímulo luminoso é um fator importante na sincronização do enxameamento, desta forma certas espécies são impelidas a subir à superfície para a reprodução durante a aurora ou crepúsculo ou ainda estimuladas pela lua, como os vermes palolo. Esses animais são bioluminescentes e a frequência e os pulsos de luz é espécie específico.

Silídeo Myrianida pachycera (Augener, 1913), brotamento de epitocos.
Em geral, após a fertilização rapidamente formam-se larvas planctônicas, larvas trocóforas. Em seguida ocorre a metamorfose que transforma a trocófora em corpo juvenil e é caracterizada pelo alongamento gradual da zona de crescimento, sendo produzidos novos segmentos nessa região. Certas espécies, entretanto, apresentam sua fase trocófora ainda dentro do ovo, eclodindo assim um indivíduo já na fase juvenil.
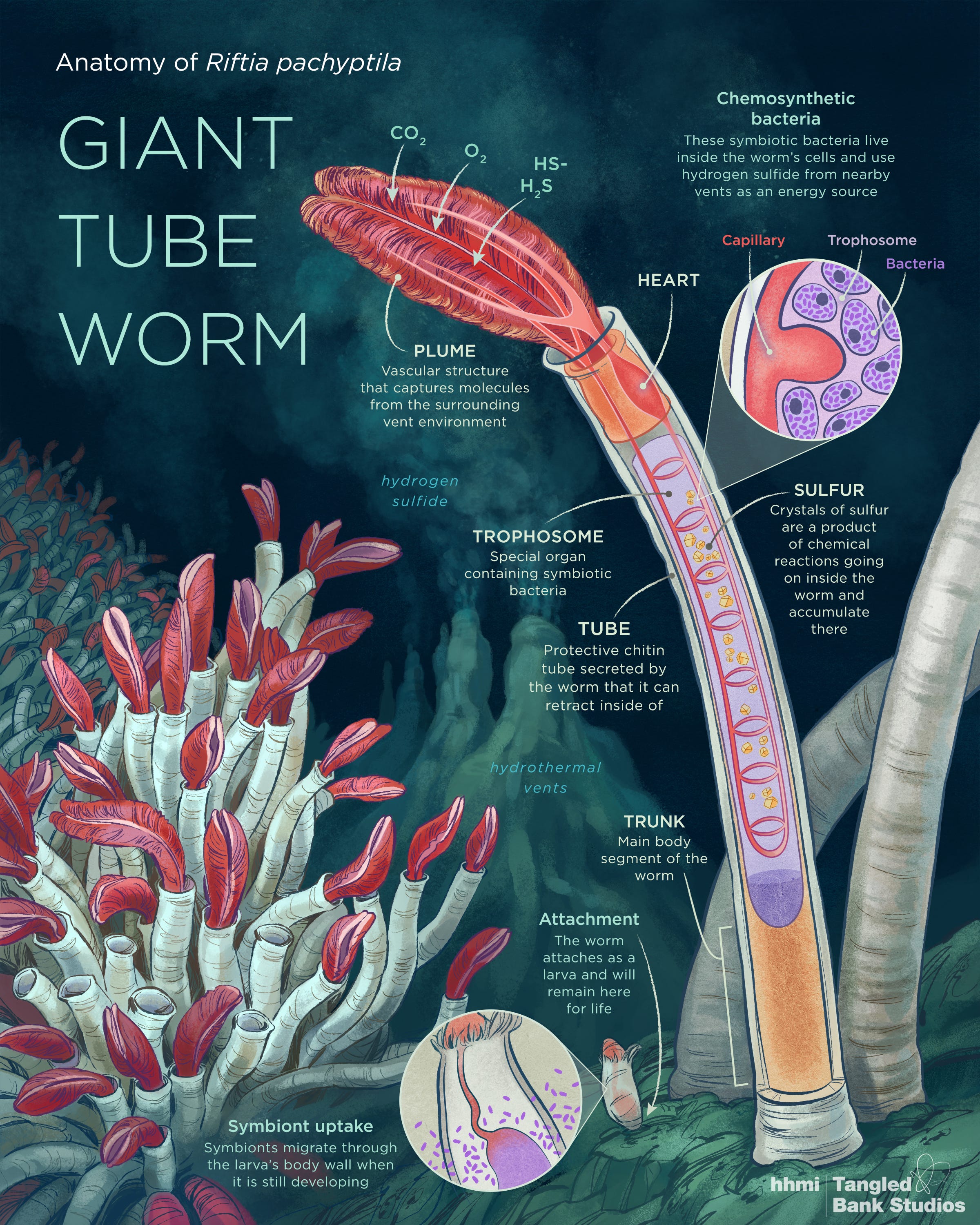
Riftia pachyptila poliqueta marinho

Poliqueta tubícola marinho de Bombinhas/SC
OLIGOCHAETA
Apresentam poucas cerdas (terrestres ex.: minhoca) do grego: ὀλίγος, oligos, pouco e χαίτη , chaite, cerda; oligos = pouco + chaeta = cerda; poucas cerdas, é uma classe dos Annelida que possui o corpo constituído de segmentos semelhantes que lembram anéis. Habitam o solo e habitats aquáticos. Comedores de depósitos não seletivo. Boca revestida internamente por cutícula, esôfago forma papo e moela, tres pares de glandulas calciferas no esofago, intestino com tiflossole, para aumento da superfície de absorção dos nutrientes. Celulas cloragógenas: detoxificação de substrancias e acumulo de lipidios; intestino posterior: absorção de água e formação das fezes. Sistema circulatório fechado, com dois vasos principais: um dorsal pulsatil (mais importante) um vaso ventral, ainda pode ocorrer um subneural; apresentam dois plexos: plexo digestório (absorção dos nutrientes) e um plexo subepidérmico (onde correm as trocas gasosas); sangue com hemoglobina, trocas gasosas ocorrem na pele (respiração cutânea). Nos cinco primeiros segmentos existem corações laterais. fecundação externa, formam casulos com gametas onde ocorre a fecundação).
O sangue flui em um sistema circulatório fechado de vasos e redes capilares. O número de vasos longitudinais principais pode variar e até cinco podem estar presentes: dorsal, ventral, subneural e dois lateroneurais. O vaso dorsal é bombeador (pulsante) e funciona como um coração, impulsionando o sangue anteriormente para dois a cinco pares de vasos circum-esofágicos, ou “corações laterais”, que ajudam a manter a pressão sanguínea e conduzem o sangue para o vaso ventral. Todos os vasos longitudinais estão interconectados em cada segmento por vasos sanguíneos adicionais. A maioria dos oligoquetos possui uma extensa rede capilar intraepidérmica. Pigmentos respiratórios, como a hemoglobina, estão geralmente presentes. As trocas gasosas realizam-se através da parede do corpo, não havendo órgãos respiratórios especializados. A rede capilar intraepidérmica provê ampla área de superfície para essas trocas gasosas.

Oligoqueta
HIRUDÍNEA
São anelídeos sem cerdas, perderam a segmentação interna, somente externamente se observam segmentos, glândulas salivares secretoras de substâncias anticoagulantes e anestésicas em espécies hematófagas, apresentam uma ventosa anterior (segmento 1-5) e uma ventosa posterior (27-33) segmento), para aderência no hospedeiro, no substrato ou para auxílio ao movimento.
As sanguessugas são mais conhecidas como hematófagas e chamam a atenção humana desde o antigo Egito.
Estes animais podem ser encontrados em pinturas parietais em templos que datam por volta de 1567-1308 a.C (Whitaker et al., 2004).
Desde então, muitas culturas têm descrito o poder medicinal das sanguessugas no tratamento de diversas doenças. O procedimento, que ficou conhecido como “blood letting”, em que hirudíneos são utilizados para drenar o acúmulo de sangue em lesões subcutâneas, ganhou atenção especial e influenciou diversas personalidades que tiveram importante papel no desenvolvimento da medicina moderna, tais como Galeno, Avicenna e muitos outros (Whitaker et al., 2004).

Uma xilogravura belga datada de 1639 representando uma mulher colocando sanguessugas em seu braço. Sobre a mesa ao lado dela está um jarro grande contendo sanguessugas. Imagem: Guillaume van den Bossche, Biblioteca Nacional de Medicina.

Uma xilogravura francesa datada de 1598 representando um cavalheiro se cobrindo de sanguessugas na tentativa de perder peso. Imagem: Pierre Boaistuau, Biblioteca Nacional de Medicina.
A terapia medicinal com sanguessugas declinou no final de 1800, para ser improvavelmente revivida na década de 1980 pelos cirurgiões plásticos como parte de um método engenhoso de tratar o problema do sangue aprisionado que envolve o tecido implantado ou reconstruído, “congestão venosa” na linguagem médica. Neste tratamento, as sanguessugas aliviam o sangue estagnado e congestionado ao redor dos tecidos implantados, evitando a coagulação e a necrose tecidual e permitindo o crescimento de minúsculos capilares preciosos em tecidos delicadamente cicatrizados. A terapia com as sanguessugas melhorou os resultados para o reimplante de amputados dedos, orelhas e dedos dos pés; para melhor cicatrização após cirurgia de reconstrução mamária; e para a sobrevivência de retalhos cutâneos comprometidos.
De fato, tem sido relatado na literatura que sanguessugas melhoram as taxas de sucesso desses procedimentos em 60 a 80%. Embora as sanguessugas tenham aplicações promissoras na cirurgia plástica moderna, seu uso também pode causar uma infecção muito rara da pele e tecidos moles devido a um clandestino que habita o intestino da sanguessuga.
Como qualquer inseto ou mamífero, o trato digestivo da sanguessuga pode abrigar uma variedade de bactérias, incluindo uma bactéria aquática conhecida como Aeromonas, que também pode ser comumente encontrada em água doce, salobra ou salgada em climas quentes. Aeromonas é um componente vital da microflora do intestino das sanguessugas, a relação simbiótica entre os dois organismos que depende da bactéria que produz nutrientes e possivelmente enzimas de digestão de proteínas e a sanguessuga que fornece glóbulos vermelhos ricos em oxigênio. A presença da bactéria, no entanto, também aumenta o risco raro de infecção durante a terapia sanguessuga.
As infecções por Aeromonas podem ocorrer 2 a 7 dias após o início da terapia com sanguessugas e podem causar feridas infectadas de gravidade variável, variando de pústulas e abscessos menores a fasceíte necrotizante e sepse. Os relatos de casos de infecção por Aeromonas após terapia com sanguessugas são esporádicos, mas estima-se que as taxas de infecção variam de 2,4% a 20%, com uma redução estimada de 30% na taxa de cirurgias bem-sucedidas. Uma infecção pós-operatória com Aeromonas pode levar amputações de dedos previamente implantados, necrose de retalhos cutâneos ou infecção sistêmica por sepse e podem exigir semanas de antibióticos intravenosos. (Fasceíte necrotizante é uma infecção que causa a a morte dos tecidos moles do corpo. É uma doença grave de aparecimento súbito que se espalha rapidamente. Os sintomas mais comuns são pele de tom vermelho ou púrpura na área afetada, dor muito intensa, febre e vômito. A fasceíte necrotizante é um tipo de gangrena).
Apesar do uso destes parasitas em tratamentos médicos ter diminuído substancialmente a partir do século XIX, devido ao avanço da medicina moderna, eles continuam sendo usados em transplantes de tecidos, reimplantação de partes amputadas e remoção de sangue estagnado (Elliot & Kutschera, 2011).
Muitas propriedades são atribuídas à secreção salivar dos hirudíneos, tais como atividades anti-inflamatória, vasodilatadora, anestésica e antibiótica (Vera, 2005), porém, a mais explorada é a propriedade anticoagulante, que já foi descrita para vários táxons . Além do uso médico e farmacêutico, as sanguessugas são importantes modelos em estudos neurológicos devido ao longo tamanho dos neurônios presentes no cordão nervoso ventral (Elliot & Kutschera, 2011) e podem ser utilizados como bioindicadores de qualidade de água em lagos, riachos e rios (Iwama, 2017). Apesar da importância histórica, ecológica e farmacêutica, há poucos estudos sobre a diversidade de sanguessugas, especialmente na região neotropical.
As relações filogenéticas entre Hirudinea e outros clitelados têm sido estudadas por diversos autores, utilizando diferentes estratégias (Iwama, 2017).
A monofilia de Clitellata é bem suportada, tanto por dados morfológicos, quanto moleculares. Contudo, as relações dentro dos grandes grupos de Hirudinea (Branchiobdellida, Acanthobdellida e Euhirudinea) continuam incertas, já que não há concordância entre as análises acerca do grupo irmão de Euhirudinea.
Euhirudinea é classicamente dividida em dois táxons, como indicado por vários autores (e.g. Pinto, 1923; Ringuelet, 1985; Sawyer, 1986; Weber, 1915).
Membros da ordem Rhyncobdellida são sanguessugas cujo parasitismo ocorre pela penetração de uma faringe protraível. Já em sanguessugas pertencentes à ordem Arhyncobdellida, há um aparato mastigatório ou a perda completa da faringe. No entanto, esta classificação mostrou-se inválida com o advento da cladística. Apakupakul et al. (1999) mostraram que a presença de faringe protraível é um caractere plesiomórfico, resultando na parafilia de Rhyncobdellida, resultado também obtido por outros autores, em estudos posteriores (Borda & Siddall, 2004). Apesar disto, os táxons pertencentes a Rhyncobdellida (Glossiphoniidae, Piscicolidae e Ozobrachidae) são monofiléticos, de acordo com análises recentes (Iwama, 2017).
A diversidade de sanguessugas conhecidas é razoavelmente bem distribuída pelo globo. De acordo com, há aproximadamente 680 espécies descritas, a maioria para águas continentais, e para a região paleártica, que é seguida em número de espécies pela região neotropical (Iwama, 2017).
Sensilas, papilas e tubérculos
A ornamentação do tegumento das sanguessugas é um importante caractere taxonômico, principalmente em grupos que apresentam uma alta complexidade na distribuição destes ornamentos. Em algumas espécies, esta ornamentação pode apresentar padrões metaméricos, que podem ser utilizados como referência para a determinação da anulação, como em membros de diversas espécies de Haementeria Filippi, 1849 (figura 1).

Figura 1. Ornamentações do tegumento de sanguessuga.
Apesar de serem conhecidas mais estruturas ornamentais em sanguessugas, as duas principais são as sensilas e os tubérculos.
As sensilas são órgãos sensoriais que podem ser utilizados para a determinação dos anéis a2, onde também está o gânglio do cordão nervoso ventral. Há muita confusão em relação a estes órgãos na literatura, Souza (1978) e Schlenz (1971) descreveram-nos como estruturas com certa elevação cônica, contrariando a maior parte da literatura (ex. Sawyer, 1986).
Esta confusão pode estar associada ao uso da expressão “papilas segmentares”, usada por Pinto (1923). Apesar de não deixar claro a natureza da estrutura estudada, Pinto (1923) relatou que estes órgãos são extremamente inconspícuos. Derosa e Friesen (1981) forneceram um estudo sistemático da morfologia das sensilas em Hirudo medicinalis, por meio de micrografia
eletrônica de transmissão. Segundo os autores, as sensilas podem ser percebidas como regiões mais claras do tegumento, que apresentam cílios, mas não há menção a qualquer elevação perceptível sob estereomicroscópio.
Estas sensilas podem, no entanto, variar de formato dependendo da região onde se encontram. Como demonstrou Elliott (1987), as sensilas localizadas próximas aos lábios da ventosa oral diferem em tamanho, distribuição e comprimento dos cílios.
Os tubérculos ou papilas, por outro lado, apresentam elevação cônica e podem apresentar seus próprios padrões de distribuição. Em membros de Haementeria, é possível determinar certos padrões metaméricos na distribuição dos tubérculos, como mostrou Souza (1978). Contudo, a morfologia dos tubérculos nunca foi estudada detalhadamente por meio de técnicas de microscopia eletrônica e estes órgãos podem facilmente ser confundidos com os órgãos de Lang, que são caracteres restritos aos anéis a2 de sanguessugas do gênero Haementeria (Oceguera-Figueroa, 2012). (Iwama, 2017).
Cada segmento se encontra externamente dividido em um número variável de anéis, sendo este número um caractere de extrema importância para a taxonomia de hirudíneos.
Gratiolet (1862) foi o primeiro autor a reconhecer o padrão metamérico de distribuição dos anéis como um importante caractere taxonômico e definiu o limite anterior do segmento como sendo o anel portador de papilas segmentares (Whitman, 1886), um conceito divergente daquele tradicional em sanguessugas, mais conhecido como Castle-Moore (Castle, 1900; Moore, 1899; 1900; Sawyer, 1986;).
Castle (1900) e Moore (1899, 1900), a partir da análise cuidadosa do padrão de anulação de sanguessugas bianuladas, concluíram independentemente que o padrão básico de anulação em Euhirudinea são três anéis por segmento e que os distintos padrões metaméricos encontrados são derivados deste básico. Os autores também propuseram que o anel portador de papilas segmentares, cuja localização coincide com a dos gânglios metaméricos do cordão nervoso ventral, correspondia àquele localizado no meio do metâmero, portanto, o segundo anel.
Baseado nesta proposição, Moore (1899) introduziu uma nomenclatura que indica a origem dos anéis. Os três anéis primários são chamados de a1, a2 e a3. Os anéis secundários (derivados diretamente dos anéis primários) recebem a conotação b1, b2, b3, b4, b5 e b6, onde cada anel, dá origem a, no máximo, dois derivados.
A figura 2a ilustra o sistema proposto por Moore (1899) e demonstra como é possível estabelecer o anel primário de origem de cada anel derivado. Apesar de útil, a conotação Castle-Moore não é totalmente adequada para representar a anelação de sanguessugas, já que é comum que espécimes apresentem padrões de anelação que não são consistentes ao redor de toda a circunferência do corpo, como mostra a figura 2D. Nestes casos, é preciso utilizar a conotação dos anéis primários para fazer referência ao conjunto de anéis derivados de tal anel primário.

Sanguessugas são frequentemente achatadas dorsoventralmente e possuem geralmente duas ventosas: uma posterior, com a qual se fixam no substrato, e outra anterior, ao redor da boca. O número de metâmeros é sempre constante em uma espécie, sendo geralmente 33.
Externamente, entretanto, hirudíneos aparentam ter um número muito maior de segmentos, porque cada um deles é marcado por sulcos transversais que formam anéis ou ânulos superficiais Outra característica marcante dos hirudíneos é seu celoma: na maioria das espécies, os septos foram perdidos, havendo fusão dos compartimentos celomáticos, além de preenchimento de grande parte do celoma por tecido conjuntivo. Os espaços celomáticos restantes estão representados por um sistema de canais (ou lacunas) que pode substituir por completo o sistema circulatório.
podem ser aquáticos e terrestres ex.: Hirudo spp. Antigamente eram chamados Achaetas, ou seja, sem cerdas. Alimentam-se geralmente do sangue de outros animais, embora muitas sanguessugas sejam predadoras carnívoras e não hematófagas. São animais hermafroditas. Celoma reduzido, gametas maturam no celoma. Fecundação interna (no celoma) e formam casulos com embriões.

Morfologia de um Hirudineo

Locomoção em Hirudínea

Movimento dos Clitellata

Hirudo spp, Hirudinea
MELHORES VIDEOS
Annelida características gerais
Poliquetas
Clitellata
Bibliografia
https://sigam.ambiente.sp.gov.br/sigam3/Repositorio/511/Documentos/APAM_LS/3.2.6.3_MEco_Bentos_APAMLS_OK.pdf
https://sigam.ambiente.sp.gov.br/sigam3/Repositorio/511/Documentos/APAM_LS/3.2.6.3_MEco_Bentos_APAMLS_OK.pdf
https://www.researchgate.net/publication/237489924_An_illustrated_key_to_the_earthworms_of_the_Samoan_Archipelago_Oligochaeta_Glossoscolecidae_Megascolecidae_Moniligastridae
https://www.fcav.unesp.br/Home/departamentos/biologia/MARIACELIAPORTELLA/5t.-aula-annelida-2014.pdf
https://www.fcav.unesp.br/Home/departamentos/biologia/MARIACELIAPORTELLA/5t.-aula-annelida-2014.pdf
https://acervodigital.ufpr.br/bitstream/handle/1884/68506/R%20-%20T%20-%20TALITA%20FERREIRA.pdf?sequence=1&isAllowed=y
http://www.sfu.ca/~fankbone/v/polychaeta.html
http://www.sfu.ca/~fankbone/v/polychaeta.html
https://www.britannica.com/animal/Protostomia
Construção de um minhocário com alunos do sétimo ano
Darwin, C. R. 1881. The formation of vegetable mould, through the action of worms. London: John Murray. Text Image PDF F1357
Darwin, C. R. 1881. The formation of vegetable mould, through the action of worms. London: John Murray. 5th thousand. Text Image F1361
Darwin, C. R. 1882. The formation of vegetable mould, through the action of worms, with observations on their habits. 6th thousand (corrected). London: John Murray. Image PDF F1362
Darwin, C. R. 1882. The formation of vegetable mould, through the action of worms, with observations on their habits. New York: D. Appleton. Image PDF F1363
Darwin, C. R. 1882. The formation of vegetable mould, through the action of worms, with observations on their habits. London: John Murray. 7th thousand. Corrected by Francis Darwin. Text Image PDF F1364.
Temir A. Britayev1, Elena Mekhova1, Yury Deart1, Daniel Martin Do syntopic host species harbour similar symbiotic communities? The case of Chaetopterus spp. (Annelida: Chaetopteridae). https://peerj.com/articles/2930/
French
Darwin, C. R. 1882. Role des vers de terre dans la formation de la terre végétale. Image PDF F1403
German
Darwin, C. R. 1882. Die Bildung der Ackererde durch die Thätigkeit der Würmer. Translated by J. V. Carus. Stuttgart: Schweizerbart. Image PDF F1404
Darwin, C. R. 1882. Role des vers de terre dans la formation de la terre végétale. Image PDF F1403
German
Darwin, C. R. 1882. Die Bildung der Ackererde durch die Thätigkeit der Würmer. Translated by J. V. Carus. Stuttgart: Schweizerbart. Image PDF F1404
(Atualizada em 17/VIII/2023)