SUPERCLASSE PISCES
PEIXES CARTILAGINOSOS
E
PEIXES ÓSSEOS

Selo comemorativo da Namibia


Megalodon

Osteichthyes
SUPERCLASSE PISCES
PEIXES
Os peixes vivem as águas salgadas dos mares e oceanos e água doce dos rios, e lagos. Provavelmente, foram os primeiros vertebrados a surgir na Terra, e eram pequenos, sem mandíbula, tinham coluna vertebral cartilaginosa e uma carapaça revestindo seus corpos.
Ao longo da evolução, houve uma série de adaptações que permitiram aos peixes melhores condições de sobrevivência em seu habitat.
Inicialmente eram filtradores de partículas nutritivas suspensas na água ou depositadas no fundo do oceano, rios e lagos. Mas esse modo de vida os tornavam presas em potencial de alguns tipos de invertebrados, ao longo de milhões de anos evoluíram características que os permitiram tornarem-se eficientes predadores.
O termo "peixe" é frequentemente utilizado para descrever um vertebrado aquático com brânquias, com forma hidrodinâmica, membros locomotores (se presentes) na forma de nadadeiras pares e ímpares, normalmente com escamas de origem dérmica (ganoides, ctenoides e cicloide) revestidas pela epiderme e com muitas glândulas de muco. Sendo este conceito do termo "peixe" utilizado por conveniência, e não como uma unidade taxonômica, porque os peixes não constituem um grupo monofilético.
Tradicionalmente os peixes são divididos em dois grupos:
CHONDRICHTHYES
Peixes com esqueleto cartilaginosos ex.: tubarões, raias e quimeras.
OSTEICHTHYES
OSTEICHTHYES
Peixes com esqueleto ósseos como por ex.: bacalhau, sardinhas, atum, salmão, garoupa.
Entretanto existem ainda outros organismos que são incluídos, para fins didáticos, nesse grupamento chamado de peixes como por exemplo, as lampreias e feiticeiras (ambos da classe Agnatha).
Entretanto existem ainda outros organismos que são incluídos, para fins didáticos, nesse grupamento chamado de peixes como por exemplo, as lampreias e feiticeiras (ambos da classe Agnatha).
RECAPITULANDO: CARACTERÍSTICAS DOS AGNATHA
- Corpo com formato cilíndrico e alongado.
- Sem mandíbula ou maxila.
- Sem escamas.
- Boca circular para sucção e dentes pontiagudos para raspar.
- Sem nadadeiras pares.
- Abas peitorais presentes em algumas formas extintas (atualmente desapareceram).
- Nas espécies ancestrais a pele era revestida por fortes escamas ósseas, que foram perdidas nas atuais.
- O esqueleto é cartilaginoso nas formas atuais e parece que nas espécies extintas elas também não eram ossificadas.
- A notocorda persiste nos adultos.
- Apresentam um "olho pineal mediano". As formas extintas e atuais apresentam uma narina única, mediana, localizada à frente do olho pineal.
- Apresentam sete ou mais aberturas brânquias.
- A faringe é utilizada na alimentação por filtração nas larvas e nos adultos das espécies extintas.
- Ex.: Lampreias e Peixe-bruxa (feiticeira)
- As feiticeiras são cavadores marinhos que fazem tocas em sedimento lodoso.
CARACTERÍSTICAS GERAIS DOS PEIXES
- Bilaterais
- Triblásticos
- Celomados, enterocelomados
- Notocorda, tubo neural dorsal oco, fendas faringeanas, coração ventral, cauda pós-anal, músculos segmentares. Ainda apresentam um epitélio de revestimento com escamas, glândulas mucosas.
- Esqueleto de origem mesodérmica que pode ser cartilaginoso ou ósseo.
- Apresentam crânio e mandíbulas.
- Apresentam temperatura variando com o meio, sendo pecilotérmicos ou heterotérmicos.
- Respiração branquial ou pulmonar.
- Podem regular sua densidade para submergir ou emergir na coluna d'água. Nos condrictes a regulação da densidade se dá pela acumulação de óleo no fígado, enquanto que nos osteíctes a regulação da densidade se da através do acúmulo de gases na vesícula natatória (bexiga natatória).
- Brânquias: livres ou não protegidas nos peixes cartilaginosos,
- Brânquias protegidas por opérculo: peixes ósseos.
- Boca: ventral ou sub-terminal nos peixes cartilaginosos e anterior ou terminal nos peixes ósseos.

Brânquias

Posição da boca

Respiração branquial em em agnatos e peixes
Evolução da respiração pulmonar em peixes
Acreditava-se que os pulmões dos peixes era uma adaptação que lhes permite viver em habitats de água doce pobres em oxigênio. No entanto, a consideração da história evolutiva do sistema respiratório do protovertebrados e dos primeiros vertebrados, o registro fóssil de peixes ósseos e a anatomia e fisiologia dos peixes pulmonados existentes podem indicar que os pulmões são uma adaptação para fornecer oxigênio ao coração (Farmer 1997, 1998, 1999). Assim, os pulmões podem ter permitido que os primeiros peixes se tornassem animais grandes e ativos em um ambiente marinho.

Esquema do sistema circulatório de larvas de lampreia e hagfish, um modelo de protovertebrado.
O sangue rico em oxigênio da respiração cutânea se mistura com o sangue pobre em oxigênio que retorna ao coração dos músculos e de outros órgãos, antes que a mistura entre no coração. Assim, o coração, que não tem circulação coronariana e depende inteiramente do oxigênio do sangue luminal (o pouco que esta ainda dissolvido no sangue), está a jusante (eferente) do órgão de troca gasosa.

Esquema do sistema circulatório de um peixe que respira guelras
À medida que os peixes ficavam maiores, a pele não era mais adequada como único "trocador" de gás. Quando as brânquias se tornaram o local das trocas gasosas, substituindo a pele, o coração foi deixado a montante (eferente) do trocador de gases. Assim, o sangue pobre em oxigênio que retorna dos músculos e de outros órgãos ao coração não é enriquecido (com oxigênio). Esses peixes podem estar limitados em sua performance ou desempenho aeróbio a uma potencial pressão seletiva para a evolução de uma circulação coronariana.

Esquema do sistema circulatório de Amia calva, um peixe que respira ar basal.
O sangue rico em oxigênio do pulmão se mistura com o sangue pobre em oxigênio que retorna ao coração vindo dos músculos e de outros órgãos, antes que a mistura entre no coração. Assim, o coração, que não tem circulação coronariana e depende inteiramente do oxigênio do sangue luminal, está a jusante (eferente) do órgão de troca gasosa. Os peixes respiradores pulmonares com este tipo de arranjo circulatório (por exemplo, o peixe pulmonar australiano, Neoceratodus forsteri, Lepisosteus sp, e Megalops sp) são peixes muito ativos e respiram ar, embora ativos independentemente da tensão de oxigênio na água.

Esquema do sistema circulatório no peixe pulmonado
sul-americano, Lepidosiren paradoxa.
Esses peixes são respiradores de ar obrigatórios. Imagine peixes que se afogam quando mantidos debaixo d'água! Eles vivem às vezes em água pobre em oxigênio e suas guelras degeneraram, especialmente os filamentos do 3º e 4º arcos branquiais, o que impede o sangue rico em oxigênio que passou pelo pulmão e flui por esses arcos branquiais de perder oxigênio para a água. O quinto e o sexto arcos branquiais recebem sangue pobre em oxigênio e rico em dióxido de carbono. Os fluxos de sangue são mantidos separados por septação do átrio em um lado direito e esquerdo, por um septo parcial no ventrículo cardíaco e por uma válvula em espiral no cone arterial. Os filamentos branquiais do 5º e 6º arcos são usados para remover o dióxido de carbono do sangue. Metade do coração carece de do sangue luminal enriquecido com oxigênio, e esses peixes não são altamente ativos.
ORIGEM
No período Cambriano, cerca de 510 milhões de anos atrás, apareceram aos primeiros tipos de peixes, denominados de ''Ostracodermos''.
Os Ostracodermos viveram nos oceanos antigos durante um curto período de tempo geológico, que durou do Cambriano (500 milhões de anos atrás) até o final do período Devoniano, cerca de 310 milhões de anos atrás.
Os Ostracodermos eram organismos muito diferentes dos peixes que hoje nós conhecemos. Eram peixes que não apresentavam mandíbulas, tendo o corpo recoberto por placas ósseas. Ostracodermos (fósseis) atualmente estão colocados na classe Agnatha ou Cyclostomata. Os Agnata atuais são peixes viventes que também não apresentam mandíbula e incluem peixes como as lampreias e feiticeiras.
CRANIATA: GNATHOSTOMATA
VERTEBRATA
A origem dos vertebrados foi definida pela evolução de um endoesqueleto de origem mesodeérmica; no entanto, pouco se sabe sobre os mecanismos de desenvolvimento responsáveis por essa inovação evolutiva marcante.
Nos vertebrados com mandíbula, a matriz da cartilagem consiste predominantemente em colágeno do tipo II (Col2α1). Recentemente foi demonstrado a presença de Col2α1 na cartilagem da lampreia, indicando que a cartilagem à base de colágeno tipo II evoluiu mais cedo do que se imaginava.
A origem da cartilagem de vertebrados tem sido investigada e foi mostrado que os peixes-bruxa, o grupo irmão das lampreias, também possui cartilagem baseada em Col2α1, sugerindo sua presença no ancestral comum dos vertebrados do grupo coroa.
Como já sabemo colágeno é o principal componente de tração do aparelho musculoesquelético, com capacidade de formar fibras com alta resistência elástica. Esta molécula compreende a maior parte da matéria orgânica da pele, tendões, ossos e dentes, desempenhando um papel estrutural nos tecidos.
Além disso o colágeno tem aplicação na engenharia de tecidos artificiais, por apresentar a função de orientar tecidos em desenvolvimento, sendo favorável a sua utilização como biomaterial. Inicialmente, o colágeno foi utilizado para produção de fios cirúrgicos devido às suas propriedades de biomaterial (baixa resposta imunológica, baixo índice de alergenicidade, biodegrabilidade e habilidade em promover crescimento celular). Atualmente utilizado na forma de membranas, tubos, pós, esponjas e soluções injetáveis. (NETO, 2011; VULCANI e PLEPIS, 2006; LEE et al., 2001).
Cefalocordado (anfioxo) ou "lancelet", um grupo irmão dos vertebrados, possui um gene ancestral do colágeno fibrilar do clado A (ColA) que é expresso na notocorda. Juntos, esses resultados sugerem que a duplicação e diversificação dos genes ColA na transição cordado-vertebrado podem ser a base da origem evolutiva dos tecidos esqueléticos dos vertebrados.
As hipóteses de relações filogenéticas dos vertebrados foram grandemente estabelecidas com base em caracteres anatômicos, particularmente aqueles do esqueleto. Os esqueletos de vertebrados com mandíbula (gnatostomados) são compostos de cartilagem e osso, que contêm altos níveis de colágeno tipo II e tipo I (Col2α1 e proteína Col1α1/2, respectivamente).
Em contraste, os esqueletos cartilaginosos de lampreias e peixes-bruxa, os únicos peixes sem mandíbula (Agnatha) existentes, tem sido relatado que não são colagenosos e contem em vez disso, as proteínas semelhantes à elastina: lamprina e mixinina.
Essa diferença nas matrizes extracelulares de esqueletos de vertebrados levou à ideia de que o colágeno tipo II se tornou o principal componente estrutural da cartilagem dos gnatóstomados após a divergência dessas duas linhagens.
No entanto, essa visão foi contestada por um trabalho recente de que as lampreias têm dois ortólogos Col2α1 e que ambos os genes são expressos durante a condrogênese.
A cartilagem de lampreia adulta também demonstrou ser rica em proteína Col2α1. Além disso, foi identificado um ortólogo de Sox9 de lampreia, um regulador transcricional direto de Col2α1 em gnatostomados, e foi mostrado que ele é co-expresso com Col2α1. Esta descoberta revelou que a via genética para a condrogênese de vertebrados antecedeu a divergência de lampreias (Agnatha) e Gnathostomata, embora precisamente quão cedo esse caráter surgiu seja desconhecido.
A origem evolutiva da cartilagem baseada em Col2α1, foi investigada; e a análise foi expandida para o clado mais inclusivo de vertebrados, que inclui os peixes-bruxa, e para um grupo irmão dos vertebrados, os anfioxos (lancelets) (Cephalochordata). Dessa forma notou-se que os peixes-bruxa (Craniata: Agnatha) também têm cartilagem baseada em Col2α1, sugerindo que esse tipo de cartilagem estava presente no ancestral comum de todos os vertebrados coroa (suggesting that this type of cartilage was present in common ancestor of all crown vertebrates.).
Uma análise de "lancelets" (anfioxo) revelou a presença de um gene ancestral do colágeno fibrilar do clado A (ColA) que é expresso na notocorda. Assim, durante a transição cordado-vertebrado, um gene ancestral ColA sofreu duplicação e diversificação, e este processo pode estar na origem da origem evolutiva dos tecidos esqueléticos de vertebrados (GuangJun Zhang and Martin J. Cohn, 2006).

Árvore de consenso estendida para a análise filogenética Bayesiana de proteínas de colágeno fibrilar do clado A. Os números em cada nó indicam valores de probabilidade posterior (pp) com base em um milhão de réplicas. Os comprimentos das ramificações são proporcionais às médias das densidades de pp para suas substituições esperadas por sítio. A árvore é enraizada pelo colágeno fibrilar do clado B do tunicado (C. intestinalis) (ColB) e colágeno fibrilar do ouriço-do-mar (ColP2α). Peixe-bruxa (Hagfish) Col2α1 (boxed) é agrupado com lampreia Col2α1a e Col2α1b com um pp de 0,99. Este clado ciclóstomo Col2α1 une-se à base do clado gnatostômio Col2α1 com um pp de 0,96. O colágeno fibrilar do clado A de anfioxo ou Lancelet (AmphiColA; em caixa) é unido à família de colágeno do clado A de vertebrados com um pp de 0,99. Os métodos de evolução mínima e de máxima verossimilhança confirmam essas posições para os peixes-bruxa Col2α1 e AmphiColA.
Extended majority-rule consensus tree for the Bayesian phylogenetic analysis of clade A fibrillar collagen proteins. Numbers at each node indicate posterior probability (pp) values based on one million replicates. Branch lengths are proportional to means of the pp densities for their expected replacements per site. The tree is rooted by tunicate (C. intestinalis) clade B fibrillar collagen (ColB) and sea urchin fibrillar collagen (ColP2α). Hagfish Col2α1 (boxed) is grouped with lamprey Col2α1a and Col2α1b with a pp of 0.99. This cyclostome Col2α1 clade joins to the base of the gnathostome Col2α1 clade with a pp of 0.96. Lancelet clade A fibrillar collagen (AmphiColA; boxed) is joined to the vertebrate clade A collagen family with a pp of 0.99. Minimum evolution and maximum likelihood methods confirm these positions for hagfish Col2α1 and AmphiColA.

Origem da cartilagem à base de colágeno tipo II. As relações filogenéticas de tunicados, lanceletes (anfioxos), peixes-bruxa, lampreias e gnatostomados (37, 38) são mostradas. As linhas tracejadas indicam posições alternativas de ramais (27). As letras apontam os principais passos na evolução da cartilagem baseada em Col2α1. O retângulo sólido indica a origem do gene Col2α1 pela(s) duplicação(ões) do gene ColA. O retângulo vazio indica a origem de Col2α1a e Col2α1b pela duplicação do gene Col2α1. (A) Expressão do gene de colágeno do clado A ancestral na notocorda de cordados-tronco. (B) Origem da cartilagem baseada em Col2α1 em vertebrados-tronco. (C) Subfuncionalização de Col2α1, em que o domínio de expressão do gene ancestral é particionado entre os genes duplicados (Col2α1a e Col2α1b); setas indicam possíveis posições deste evento.
Origin of type II collagen-based cartilage. Phylogenetic relationships of tunicates, lancelets, hagfishes, lampreys, and gnathostomes (37, 38) are shown. Dashed lines indicate alternative branch positions (27). Letters point out major steps in Col2α1-based cartilage evolution. The solid rectangle indicates the origin of the Col2α1 gene by ColA gene duplication(s). The hollow rectangle indicates the origin of Col2α1a and Col2α1b by Col2α1 gene duplication. (A) Ancestral clade A collagen gene expression in notochord of stem chordates. (B) Origin of Col2α1-based cartilage in stem vertebrates. (C) Subfunctionalization of Col2α1, in which the expression domain of the ancestral gene is partitioned between the duplicate genes (Col2α1a and Col2α1b); arrows indicate possible positions of this event.
O grupo fóssil Placodermi é o mais filogeneticamente basal do clado de vertebrados com mandíbulas, mas carece de uma dentição marginal comparável à dos possuidores de dentes: Chondrichthyes, Acanthodii e Osteichthyes (Gnathostomata do grupo da coroa). Os dentes dos gnatostomados do grupo coroa são parte de uma dentição ordenada substituída e padronizada por uma lâmina dentária, exemplificada pelo modelo elasmobrânquico. Uma dentição reconhecida por esses critérios foi previamente considerada ausente em placodermes, com base em evidências estruturais, como ausência de verticilos e dentina típica de vertebrados.
Os Gnathostomata, ou gnatostomados, (animais com mandíbula) em sua maioria são do Devoniano Médio (380 milhões de anos atrás) ao presente. Eles diferem de todos os outros cordados que os precederam (Cefalocordados, urocordados e craniados agnata) por apresentarem um dispositivo de mordedura vertical, as mandíbulas, que se originaram de arcos branquiais.
Além da mandíbula, esses animais apresentam órgãos de apreensão, corte, esmagamento ou cisalhamento, ou seja, os dentes.
Entre os vertebrados recentes, os primeiros gnatostomados incluem tubarões, raias, quimeras, peixes com nadadeiras raiadas, e peixes com nadadeiras lobadas. Outro grupo de gnatostomados iniciou a exploração do ambiente terrestre este inclui os anfíbios os quais deram origem aos tetrápodes.

Origem embrionária do crânio ósseo em cinco organismos modelo de vertebrados dispostos em uma filogenia simplificada de vertebrados. Territórios derivados da crista neural (azul) foram verificados experimentalmente em cada espécie, embora as contribuições específicas de fluxos migratórios (de células) individuais sejam relatadas apenas para galinhas, axolotl e Xenopus. A derivação de componentes restantes do mesoderma (magenta) foi verificada experimentalmente em camundongos e galinhas e é presumida para as espécies restantes. As pontas de seta apontam para a interface da crista neural-mesoderma no teto do crânio, que está deslocada caudalmente em Xenopus. Os dados para peixe-zebra são das referências 8, 9; o diagrama é baseado na ref. 8 (figura reproduzida com permissão da PLoS). Os dados para axolotl e Xenopus são do presente estudo; os crânios são redesenhados a partir das referências 16, 42, respectivamente (figuras reproduzidas com permissão de John Wiley and Sons). Os dados para frango são da ref. 43; o diagrama é baseado na ref. 44 (figura reproduzida com permissão de John Wiley and Sons). Os dados para mouse são das referências 7, 45; o diagrama é baseado nas referências 4, 46 (figura reproduzida com permissão de John Wiley and Sons). F, frontal; Fp, frontoparietal; N, nasal; P, parietal; Px, pré-maxila; Quadrado, esquamosal.
(Embryonic origin of the bony skull in five vertebrate model organisms arrayed on a simplified vertebrate phylogeny. Neural crest-derived territories (blue) have been verified experimentally in each species, although the specific contributions from individual migratory streams are reported only for chicken, axolotl and Xenopus. Derivation of remaining components from mesoderm (magenta) has been verified experimentally in mouse and chicken and is presumed for the remaining species. Arrowheads point to the neural crest–mesoderm interface in the skull roof, which is displaced caudally in Xenopus. Data for zebrafish are from refs 8, 9; diagram is based on ref. 8 (figure reproduced with permission from PLoS). Data for axolotl and Xenopus are from the present study; skulls are redrawn from refs 16, 42, respectively (figures reproduced with permission from John Wiley and Sons). Data for chicken are from ref. 43; diagram is based on ref. 44 (figure reproduced with permission from John Wiley and Sons). Data for mouse are from refs 7, 45; diagram is based on refs 4, 46 (figure reproduced with permission from John Wiley and Sons). F, frontal; Fp, frontoparietal; N, nasal; P, parietal; Px, premaxilla; Sq, squamosal. Fonte).
Superclasse Pisces
Chondricthyes e Osteichthyes
Dentro dos gnatostomados mais basais encontramos um grande grupo que constitui a superclasse Pisces. Esta superclasse divide-se em dois clados principais, os Chondrichthyes e Osteichthyes.

Coluna de tempo geológico
Os primeiros fósseis de vertebrados semelhantes a peixes são fragmentários demais para permitir rastrear os peixes modernos precisamente até suas origens. Acredita-se, baseado nos fósseis até agora conhecidos que as formas ancestrais evoluíram no final do Período Ordoviciano (cerca de 455 milhões de anos atrás) nas partes superiores de rios. Durante o final do Siluriano e o início Devoniano e dos períodos que se seguiram, apareceu um grupo extremamente diverso de peixes com uma armadura de placas ósseas, com estruturas semelhantes a mandíbulas, nadadeiras pares e tecido ósseo esquelético.
Os paleontólogos referem-se a essas formas extintas como uma classe distinta, Placodermi.
Entre o início e o fim do Devoniano (o último há cerca de 350 milhões de anos), os placodermes atingiram seu auge em diversidade e número e quase desapareceram completamente; apenas alguns permaneceram outros 10 milhões de anos seguintes no subperíodo do Mississippiano (aproximadamente, o Carbonífero Inferior).

Chondrichthyes: Cladoselache e Hybodus dois peixes cartilaginosos extintos.
(Fonte: Encyclopædia Britannica, Inc.)
Embora algumas formas semelhantes a tubarões tenham permanecido em ambientes de água doce, a grande maioria logo invadiu o oceano, talvez em resposta ao árido clima devoniano. Lá, eles se adaptaram à vida na água salgada, desenvolvendo o hábito de retenção de uréia. Seu esqueleto cartilaginoso, longe de representar um estágio evolutivo anterior aos Osteichthyes, como se acreditava, é mais do que provável degenerado em vez de primitivo. Possivelmente, seus precursores foram os petalichthyids, um grupo de placodermos parecidos com tubarões do Devoniano, que tinham esqueletos ossificados e nadadeiras bem desenvolvidas.
A relação filética das quimeras e dos tubarões e raias é um assunto aberto a várias interpretações. Embora ambos os grupos tenham muitas características em comum (como a posse de um esqueleto cartilaginoso, escamas placóides, dentes simplesmente embutidos na gengiva, válvula em espiral no intestino, hábito de retenção de uréia, fertilização interna (para a qual os machos têm clásper) e ausência de bexiga natatória), os dois grupos podem ter evoluído independentemente ao longo de linhas evolutivas paralelas. As quimeras evoluíram a partir dos picodontes, uma ordem de placodermos devonianos com forma corporal e estrutura dentária muito sugestiva das quimeras modernas.

Comparação entre os pesos pesados cartilaginosos


Tamanho dos condríctes atuais e extintos

Diversidade de espécies de raias e tubarões existentes nas Américas. O número total de espécies é baseado no site FishBase (Froese & Pauly, 2017), Ocean Biogeographic Information System (OBIS: http: // www.iobis.org/), Tabela 1; Dados S1; Tabela S2. Tamanho real DOI: 10.7717 / peerj.5313 / fig-1.
(Fonte Ocean Biovidiv. Inform. Sys.).

Mapa mostrando a distribuição das espécies de tubarões e raias mais comumente observadas e registradas em levantamentos de recifes rochosos em Galápagos divididas em biorregiões, conforme proposto por Edgar et al. (2004). As biorregiões de Cold West e Elizabeth foram fundidas, pois os dados da biorregião de Elizabeth eram escassos. Os ícones (imagens, silhuetas) das espécies não estão apresentados em escala.
Map showing the distribution of the sharks and rays species most commonly observed and recorded in rocky reef surveys in Galapagos divided in bioregions, as proposed by Edgar et al. (2004). The bioregions of Cold West and Elizabeth have been fused, as data from Elizabeth bioregion was scarce. The species icons are not presented in scale. (Fonte: Elasmobranchs of Galapagos marine reserve)

Diversidade de Chondrichthyes existentes

Modelos de silhueta visualizando as dimensões do corpo de †Otodus megalodon com base em nossas extrapolações em comprimentos totais diferentes.
(a) ~16 m, (b) ~3 me (c) ~8 m. Abreviações como na Tabela 1. Silhuetas criadas no Adobe Illustrator CC 2018. (d) Reconstrução paleoartística de um † O. megalodon de 16m, contrastado com um humano de 1,65 m (ilustração de Oliver E. Demuth). As formas das barbatanas são baseadas em nossas formas morfométricas generalizadas nas silhuetas, enquanto a reconstrução visa capturar suas verdadeiras formas biológicas, ou seja, a ceratotriquia da cauda presente em todos os cinco análogos modernos. (Body dimensions of the extinct giant shark Otodus megalodon: a 2D reconstruction Sep, 03. 2020)
Sinapomorfias
1. Calcificação prismática da cartilagem

Em espécies que nadam com grande velocidade esses dentículos recobrem a cavidade oral, nessas espécies mais velozes os dentículos apresentam quilhas para uma melhor perfomance no nado.

Dentículos dérmicos de um tubarão veloz.
3. Mixopterígio ou clásper
Órgão presente nos machos para cópula. São estruturas pares associados às nadadeiras pélvicas dos machos, e são estruturas complexas com várias cartilagens articuladas usadas para transferir o esperma para a cloaca das fêmeas.

Características dos condrictes
Os elasmobrânquios (raias, tubarões e quimeras) são vertebrados pertencentes à Classe Chondrichthyes, e que exibem um conjunto de características únicas dentre os Craniata; Gnathostomata (vertebrados), compondo um grupo monofilético dividido em duas subclasses distintas:
Elasmobranchii e Holocephali (COMPAGNO, 1999).
Esqueleto cartilaginoso
Os Chondrichthyes são caracterizados por um tipo especial de tecido duro que reveste as cartilagens do endoesqueleto: a cartilagem prismática calcificada.
O endosqueleto cartilaginoso de peixes Chondrichthyes (tubarões, raias e quimeras) exibe arranjos complexos e morfologias de tecidos calcificados que variam com a idade, espécie, comportamento alimentar e localização no corpo.
A cartilagem dos peixes cartilaginosos pode ser areolar, globular e prismática. O esqueleto tesselado é composto por pequenos blocos (tésselas) de cartilagem calcificada (tanto prismática quanto globular) cobrindo um núcleo de cartilagem não mineralizada, enquanto a cartilagem vertebral geralmente contém todos os três tipos de calcificação.



Crânio de um tubarão martelo ou cação martelo

Cartilagens e músculos do crâneo

Diferentes tipos de cartilagens ocorrem nos tubarões, raias e quimeras

Cargilagens do esqueleto
Presença de escamas placoides



Escamas placoides
As escamas do tipo placoide são exclusivas dos condrictes. São semelhantes a dentes pequeninos, formados por dentina e revestida por esmalte (esmalte é um tecido altamente mineralizado, muito duro), imersas no corpo do animal por uma base. Quando passamos a mão na pele de um tubarão no sentido da cauda para a cabeça a ponta da escama pode esfolar nossa pele, conferindo uma textura áspera semelhante a uma lixa. Este tipo de escama reduz enormemente o arrasto da água (devido a viscosidade desse líquido) sobre o corpo do peixe, possibilitando um deslocamento sem muito gasto de energia. Cada uma das escamas e os vales que se formam entre elas dissipam o arraste ("atrito") do corpo com a água.

Outline of a representative shortfin mako shark Isurus oxyrinchus showing the 16 regions sampled for scale photography, angle measurement, and histology. The mean erection angle for five scales in each region is given beside the nomenclature for each region. Flank scales (B2, B5, A2) are capable of greater erection angles than dorsal (B1, B4, A1) and ventral (B3, B6, A3) regions. All regions on the pectoral fin (P1, P2, P3) differed in the degree of erection possible with the leading edge scales (P1) incapable of erection. Erection angle did not differ among the three regions on the caudal fin (C1, C2, C3).

À esquerda, Micrografias eletrônicas de varredura de escamas placóides (dentículos dérmicos) das regiões dorsal, lateral e ventral do corpo (de um tubarão) entre a nadadeira peitoral e a dorsal (B1, B2, B3 na Fig. 1), de uma fêmea TL de 192 cm shorfin mako shark Isurus oxyrinchus. Três riblets (costeletas) ou quilhas são visíveis em cada escama. Anterior está para a esquerda, embora as escamas placóides nessas fotos não estejam exatamente orientadas para o eixo longitudinal do tubarão. A forma da escama placóide varia entre as regiões do corpo. (À direita) Seção sagital através de escamas placóides das regiões dorsal, lateral e ventral do corpo entre a barbatana peitoral e a barbatana dorsal (B1, B2, B3 na Fig. 1) de um tubarão mako macho de 158 cm TL (158 cm TL male shortfin mako shark). As bases das escamas (B) estão ancoradas na derme (D), e a fina epiderme (E) é visível entre as escamas. A cavidade da polpa ( ou pulpar) (PC) é visível abaixo da coroa (CR) em algumas seções. A região anterior está à esquerda.
(Left) Scanning electron micrographs of placoid scales (dermal denticles) from the dorsal, lateral, and ventral regions of the body between the pectoral fin and dorsal fin (B1, B2, B3 in Fig. 1), of a 192 cm TL female shortfin mako shark Isurus oxyrinchus. Three riblets or keels are visible on each scale. Anterior is to the left although the placoid scales in these pictures are not exactly oriented to the longitudinal axis of the shark. Placoid scale shape varies among regions of the body. (Right) Sagittal section through placoid scales from the dorsal, lateral, and ventral regions of the body between the pectoral fin and dorsal fin (B1, B2, B3 in Fig. 1) of a 158 cm TL male shortfin mako shark. The bases of the scales (B) are anchored in the dermis (D), and the thin epidermis (E) is visible between the scales. The pulp cavity (PC) is visible below the crown (CR) in some sections. Anterior is to the left. (Lang et all, 2012)

Corte sagital através de escamas placóides das regiões de borda de ataque, meio da nadadeira e borda de fuga da nadadeira peitoral (P1, P2, P3 na Fig. 1) de um tubarão mako macho TL de 158 cm. As bases das escamas (B) estão ancoradas na derme (D), e a epiderme fina (E) é visível entre as escamas. A cavidade pulpar (PC) é visível em algumas das escamas, e nem todas as escamas são seccionadas pelo centro, resultando em alguns comprimentos de coroa (CR) parecendo mais curtos do que outros. Os raios da barbatana Ceratotrichia (CE) são visíveis na região P3 porque esta parte da barbatana é muito fina. A região anterior do animal é para a esquerda, (como podemos perceber pela oritenação das escamas).
Sagittal section through placoid scales from the leading edge, mid-fin, and trailing edge regions of the pectoral fin (P 1, P 2, P 3 in Fig. 1) of a 158 cm TL male shortfin mako shark. The bases of the scales (B) are anchored in the dermis (D), and the thin epidermis (E) is visible between the scales. The pulp cavity (PC) is visible in some of the scales, and not all scales are sectioned through their center, resulting in some crown (CR) lengths appearing shorter than others. Ceratotrichia (CE) fin rays are visible in region P 3 because this part of the fin is very thin. Anterior is to the left. (Shark sin boundary layer control. Lang et all, 2012).

https://youtu.be/EP8MjFiUSbM
Shark skin equação

Os Chondrichthyes incluem dois principais clados existentes:
os Elasmobranchii e os Holocephali. (ver Biogeographic pattern in the cartilagenous fauna (Pisces: Elasmobranchii e Holocephalii in the southwest pacific ocean)

Ancestral agnata dos gnatostomados

Evolução dos gnatostomados até os peixes cartilaginosos
Alguns exemplos de peixes cartilaginosos






Quimeras

Estrutura de uma quimera



Subclasse Holocephali (quimeras, tubarões fantasmas)
Esqueleto cartilaginoso, 4 pares de brânquias, coberto em cada lado do corpo por uma prega opercular de pele levando a uma única abertura branquial externa. Primeira barbatana dorsal e espinhal (segunda nadadeira dorsal) erétil. Pele com pequenos dentículos ao longo da linha média das costas em algumas espécies e nos tentáculos e clásper dos machos. Dentes unidos para formar placas de moagem. Os cláspers dos machos são complementados por um órgão erétil, chamado tentáculo, na frente das nadadeiras pélvicas, e todos, exceto 1 gênero (Harriotta), têm outro tentáculo em forma de taco na testa. Ovos ovíparos, elípticos, em forma de fuso ou girino, envoltos em cápsulas córneas marrons, notavelmente grandes em proporção ao tamanho do pai. Na respiração, os quimeróides absorvem água principalmente pelas narinas e daí por meio de ranhuras que conduzem à boca, que geralmente é mantida fechada. Distribuído de forma variada nas zonas temperadas e boreais de todos os oceanos, nas águas costeiras e nos estuários dos rios e no mar até mais de 2.500 metros. Devoniano tardio até o presente.

Filogenia dos Cordados. Forma do corpo: o corpo é fusiforme, hidrodinâmico, comprimido (achatado látero-lateralmente) ou deprimido (achatado dorso-ventralmente).

Forma do corpo e nadadeiras ou barbatanas

Circulação sanguínea
O sistema circulatório nos peixes é fechado (sangue circula dentro de vasos) e apresenta um coração muscular com duas cavidades um átrio e um ventrículo.
Neste tipo de circulação todo o percurso do sangue é realizado dentro dos vasos sanguíneos. É mais evoluída que a circulação simples, o coração é mais musculoso, há capilares, a pressão sanguínea e velocidade do fluxo são maiores e a quantidade de nutrientes que pode ser transportado por unidade de tempo também é maior.



Circulação fechada simples
O coração dos peixes tem duas cavidades um átrio e um ventrículo, e por ele circula apenas sangue venoso, não-oxigenado. Depois de passar pelo coração, o sangue não oxigenado vai para uma artéria e dai para as brânquias, onde ocorrem as trocas gasosas. O gás carbônico é liberado na água que passa pelos filamentos branquiais e o oxigênio da água é capturado. A seguir, esse sangue, agora sangue arterial, oxigenado, é distribuído para todos os órgãos do corpo do animal. Ao voltar para o coração, está carregado com gás carbônico novamente.
Pode-se dizer, então, que a circulação é simples e completa, ou seja, o sangue passa uma única vez pelo coração para completar o ciclo e não há mistura de sangue venoso com arterial.
Reprodução
São animais dióicos (sexos separados), reprodução sexuada e fecundação interna.

https://youtu.be/TWBLi1M8pGg
Dimorfismo sexual em peixes cartilaginosos
Os tubarões podem reproduzir-se de três maneiras diferentes, podendo ser ovíparas, vivíparas, e ovovivíparas. Seja qual for o tipo de reprodução, as crias quando nascem tornam-se, automaticamente, independentes da sua progenitora.
A reprodução dos peixes cartilaginosos ocorre por fecundação interna como dito acima, na qual o macho introduz o órgão copulador masculino chamado clásper pélvico ou mixopterígio que é uma estrutura derivada da parte posterior da nadadeira pélvica (barbatana pélvica), formando o órgão copulador masculino o qual é introduzido no órgão copulador feminino (oviduto) da fêmea. Clásper são dois órgãos alongados e cônicos com o formato de duas pequenas nadadeiras que funcionam como um auxiliador na cópula.




Mixopterígio ou Clásper




https://youtu.be/vIp8rGmYkYI
Cópula de tubarão galha preta do recife
https://youtu.be/kUVp8kRWG3M
https://youtu.be/vIp8rGmYkYI
Blacktip Reef sharks mating in lagoon at Misool Eco Resort, the males had been chasing this female, bumping into her, biting her and fighting each other for quite a while before one lucky male got to mate with her.

Reprodução em peixes cartilaginosos
Nas espécies ovíparas, após a cópula onde ocorre a fecundação interna as fêmeas produzem ovos geralmente com uma casca grossa e coriácea com ganchos que se prendem em algas ou a outros substratos submersos. Desenvolvimento direto sem estágio larval. As espécies ovíparas representam cerca de 20% dos tubarões e produzem ovos retangulares, ou cápsulas, dentro dos quais se desenvolvem as crias. Os ovos são protegidos por uma membrana filamentosa acastanhada que ajudará a sua fixação nos substratos facilitando também a sua libertação através da progenitora.

Scyliorhinus canicula

Ovos de Scyliorhinus sp em diferentes estágios de desenvolvimento

Scyliorhinus sp


Embrião retirado do ovo
Os tubarões do gênero Scyliorhinus são encontrados na plataforma continental e talude de regiões tropicais a temperadas, em profundidades de 50 a 550m. São animais de pequeno a médio porte (a maioria inferior a 80cm de comprimento total). Alimentam-se de peixes, lulas e crustáceos e pela baixa motilidade são adequados para manutenção em cativeiro. Reproduzem-se por oviparidade, depositando seus ovos geralmente em pares, com período de incubação variando de 8 a 12 meses (Paiva et alli, 2015).
Nas espécies vivíparas (algumas poucas espécies), os embriões desenvolvem-se totalmente dentro do corpo da fêmea, alimentando-se de substâncias que retiram do sangue materno por meio de uma estrutura equivalente a uma placenta. O tempo de gestação é bastante longo, podendo atingir os dois anos. O número de crias varia bastante podendo ir de um (tubarão tigre da areia) a cerca de trezentos (tubarão-baleia). Os tubarões vivíparos têm no útero, uma ou mais placentas dentro das quais se desenvolvem os filhotes. Os filhotes nascem já completamente formados, e rapidamente se tornam independentes.


Enquanto nas espécies ovovivíparas as fêmeas retêm os ovos no interor do corpo até o final do desenvolvimento embrionário, dando à luz jovens imaturos.
As espécies vivíparas representam cerca de 10% dos tubarões e existe desenvolvimento direto do embrião dentro do útero materno com ligações placentárias e cordão umbilical. Quando as crias já estão completamente desenvolvidas são libertadas para o exterior.
As espécies ovovivíparas representam cerca de 70 % dos tubarões. Os embriões tem desenvolvimento direto dentro do útero materno. Usando apenas as reservas do saco vitelínico. Quando os embriões atingem o tamanho ideal são libertados para o exterior. Nas espécies ovovivíparas ocorre um fenômeno peculliar: o embrião mais forte acaba devorando os outros, essa característica é chamada de canibalismo embriônico intrauterino, nascendo apenas um filhote, o maior.
https://youtu.be/ZrqgPjZ07Ts
Canibalismo embrionário intrauterino
https://youtu.be/BWI60arRyG4
Resumo
A maioria das espécies de tubarões são ovovivíparas. Na terminologia básica, isso significa que, após a fertilização, os ovos são formados e chocados dentro da mãe antes que os filhotes vivos nasçam na água circundante. Durante o acasalamento, o tubarão macho insere o clásper (semelhante ao pênis de mamífero), (entretanto esse órgão não é homólogo ao pênis dos mamífero, i.e. não tem a mesma origem embrionária) no oviduto da fêmea e libera esperma. Esses gametas masculinos (espermatozóides) fertilizam vários de seus óvulos. Neste ponto, alguns tubarões põem seus ovos (conhecido como oviparidade), nutrem esses filhotes com uma placenta (chamada viviparidade) ou, como é o caso das espécies ovovivíparas, chocam os ovos dentro do oviduto antes de dar à luz algum tempo mais tarde; tempo que varia de espécie para espécie. Os filhotes de espécies ovovíparas são sustentados e alimentados pelo vitelo, gema dos óvulos não fertilizados, bem como por secreções especiais das glândulas nas paredes do oviduto. Depois que os filhotes se desenvolvem suficientemente dentro dos ovos, eles eclodem no oviduto. Isso geralmente ocorre após aproximadamente três meses. Eles continuam a se alimentar das gemas dos óvulos restante, bem como das secreções das glândulas, em alguns casos pode ocorrer canibalismo embriônico antes do nascimento (canibalismo embriônico intrauterino).
Excreção e equilíbrio osmótico
Regulação osmótica e iônica em peixes de água doce e salgada.
Peixe de água salgada
Problema perda de água por osmose
Resposta:
Células branquiais que transportam ativamente sais do sangue para a água do mar.
Glomérulos de Malpighi pequenos para reduzir a filtração de água e transporte ativo de sais do meio interno para o tubo urinífero, donde resulta urina concentrada.
Peixe de água doce
Problema entrada de água por osmose
Resposta:
células branquiais que transportam ativamente sais da água para o sangue.
Glomérulos de Malpighi grandes para aumentar a filtração de água e transporte ativo de sais do tubo urinífero para o meio interno, donde resulta urina diluída.
A maioria dos vertebrados marinhos mantém concentrações mais baixas de sais e outros produtos químicos em seu sangue do que as encontradas na água do mar. Como resultado, esses animais enfrentam um problema contínuo de perda de água para o meio ambiente, devido à tendência da água se mover através das membranas de regiões de baixa concentração de sal para regiões de maior concentração.
Os peixes cartilaginosos marinhos diferem de quase todos os peixes ósseos (exceto os celacantos e peixes pulmonados estivantes (torpor prolongado ou dormência de um animal durante um período quente ou seco) por serem capazes de reabsorver nos túbulos renais (rins) a maior parte de seus resíduos nitrogenados (ureia e óxido de trimetilamina) e acumular esses produtos em seus tecidos e sangue, uma capacidade denominada hábito de retenção de uréia.
A concentração dentro do corpo, portanto, excede a da água do mar circundante, e a água se move para o corpo sem nenhum gasto de energia. Quando qualquer um desses peixes mudar da água salgada para água doce, como muitos fazem, o fluxo de urina para o exterior aumenta; conseqüentemente, a concentração de ureia no sangue diminui.
Elasmobrânquios puramente de água doce, como as arraias dos sistemas de drenagem do Orinoco e do Amazonas, parecem não ter o hábito de retenção de ureia.

Regulação osmótica e iônica em peixes de água doce e salgada.
A: em água doce, os peixes perdem sais (NaCl) por difusão e ganham água por osmose (setas abertas). O transporte ativo de eletrólitos (setas preenchidas) na guelra e nos rins serve para recuperar o sal e excretar água. B: na água do mar, os peixes ganham sais (NaCl) por difusão e perdem água por osmose. Os mecanismos de transporte ativos na brânquia agora secretam NaCl e o rim excreta Mg, SO4 e outros íons divalentes. Além dos mecanismos branquiais e renais da homeostase do fluido extracelular, a tolerância das pressões osmóticas plasmáticas variando de 280 mosmol/kgH2O em água doce a 350 mosmol/kgH2O em água do mar ajuda a reduzir os desafios ambientais de sal e água.
(Osmotic and ionic regulation in freshwater and seawater fish. A: in freshwater, fish lose salts (NaCl) by diffusion and gain water by osmosis (open arrows). Active transport of electrolytes (filled arrows) in the gill and kidney serve to recover salt and to excrete water. B: in seawater, fish gain salts (NaCl) by diffusion and lose water by osmosis. Active transport mechanisms in the gill now secrete NaCl, and the kidney excretes Mg, SO4, and other divalent ions. In addition to the branchial and renal mechanisms of extracellular fluid homeostasis, the tolerance of plasma osmotic pressures ranging from 280 mosmol/kgH2O in freshwater to 350 mosmol/kgH2O in seawater helps reduce environmental salt and water challenges).
Os peixes marinhos excretam muito pouca urina, De fato alguns peixes de água salgada nem têm glomérulos nos rins.
Enquanto os peixes de água doce e água salgada enfrentam gradientes osmóticos semelhantes (300-400 mOsmol) entre seu sangue e o ambiente, a atividade de ATPase nas brânquias do peixe água salgada é relativamente maior. A razão para isso é que o sal deve ser bombeado ativamente para o peixe no intestino para que a hidratação ocorra. Assim, nas brânquias, os peixes água salgada devem livrar seu corpo do sal adquirido ativa e passivamente. Além disso, pode ser necessário para os peixes água salgada empregar bombeamento de ATPase de Na+ em troca de NH4+ para se livrar de resíduos nitrogenados, às vezes. Obviamente, Na+ -NH4+ ATPase e Cl- HCO3-ATPase não vão ajudar na osmorregulação em peixes marinhos como se presume que façam em peixes de água doce.
Tipos de rins



Os órgãos excretores consistem em rins, ureteres e bexiga urinária. A bexiga urinária não é homóloga à dos vertebrados superiores.
Rim: Os rins dos vertebrados são constituídos por néfrons ou túbulos renais. Em vertebrados ancestrais, o rim possui um néfron para cada um dos segmentos corporais que ficam entre as extremidades anterior e posterior do celoma.
O néfron é drenado para um ducto denominado ducto de Wolff ou arquinéfro localizado posteriormente à cloaca. Esse tipo de rim é conhecido como holonéfro porque se estende por todo o comprimento do corpo.
O holonéfros é encontrado hoje nas larvas de certos ciclostomos, mas não em qualquer adulto.
Em peixes e anfíbios, os túbulos mais anteriores foram perdidos, alguns dos túbulos médios estão associados a testes (testículos) e há uma concentração e multiplicação dos túbulos posteriormente. Esse rim é conhecido como rim posterior ou opistonefros.
Geralmente em peixes, os túbulos da região anterior tornam-se funcionais no início da vida e são designados como pronéfros. E os túbulos presentes nas regiões posteriores assumem função excretora ao longo da vida. Esta região dos túbulos funcionais é conhecida como mesonefros.
Os rins ocupam posição dorsal na cavidade do corpo e são situados logo ventralmente à coluna vertebral. Nos teleósteos, o rim é distinguido em regiões da cabeça e do tronco, claramente discerníveis nas carpas.
O rim da cabeça é não excretor e tem função endócrina, enquanto o rim do tronco (rim posterior) é de natureza excretora.
As peculiaridades são mencionadas a seguir:
1. O rim da cabeça é de natureza endócrina. Possui glândula inter-renal homóloga ao córtex adrenal de mamíferos.
Possui células cromafins, também semelhantes à medula adrenal dos mamíferos. 2. Nos rins estão incorporados corpos amarelos chamados corpúsculo de Stannius. É endócrino em função.
3. O rim da cabeça é o local para o desenvolvimento do sangue.
4. Ambos rins da cabeça e do tórax contém folículos heterotróficos da tireoide.
Trocas gasosas
Os tubarões respiram principalmente abrindo a boca enquanto expandem a cavidade bucofaríngea e contraem as bolsas branquiais para fechar as fendas branquiais. Com a boca fechada, eles contraem a cavidade bucofaríngea enquanto dilatam as bolsas branquiais, puxando a água sobre as brânquias onde ocorre a troca de oxigênio e gás carbônico. Então, com a boca ainda fechada, eles contraem a cavidade bucofaríngea e as bolsas branquiais, e as fendas branquiais são abertas para expelir a água.
Sistema nervoso e órgãos dos sentidos
A maioria deles ouve mal e outros enxergam mal, como o tubarão por exemplo, em circunstâncias normais, tem a visão adaptada apenas para enxergar objetos próximos, ou seja, é míope (segundo critérios humanos). Mas, em compensação, os peixes os peixes possuem um sistema sensorial exclusivo, ausente em praticamente todos os mamíferos. Esse sistema chama-se linha lateral, que recebe esse nome por estar localizada na lateral do corpo. Esse órgão forma uma linha sob as escamas do peixe.

Cérebr de um tubarão baleia

Cérebro de três especies de tubraões
Telencéfalo – Memória e função cognitiva
Lobo óptico – Interpreta as informações da visão
Nervo óptico – Leva os estímulos visuais para o lobo óptico
Cerebelo – Centro de coordenação muscular e equilíbrio
Corda espinhal – Leva e trás informações para o resto do corpo (SNP)
Hipotálamo – Secreta hormônios que estimulam a hipófise e regula funções como sono, fome.
Hipófise – Importante reguladora do metabolismo, reprodução.
Sistema hipotálamo hipófise em peixes: esta dividido em três áreas principais: Hipotálamo (parte do diencéfalo)
Neurohipófise
Adenohipófise (pró, meso e meta)
Funções do sistema hipotálamo hipófise:
Manutenção das rotas hormonais
Sintese de neuro-hormonios pelo hipotálamo
Estímulo da hipófise para liberar hormônios tróficos
Estimulo de glândulas e órgãos


Sistema da Linha Lateral em tubarrões
(Fonte: A shark's 6th sense)

https://youtu.be/9S8a5hSc22s

Ampolas de Lorenzini

Ampolas de Lorenzini



Localização da linha lateral
https://youtu.be/JDPFR6n8tAQ
How Do Sharks and Rays Use Electricity to Find Hidden Prey?
https://youtu.be/B1np7R33D1k
Muitos peixes têm a capacidade de perceber campos elétricos e produzir campos e descargas elétricas. No entanto os Elasmobranchii (tubarões e raias) apresentam o sentido de percepção de campos elétricos muito desenvolvido.
E isto se deve ao fato de que esses peixes apresentam estruturas especializadas denominadas ampolas de Lorenzine. Estas estruturas encontram-se na cabeça dos tubarões e nas nadadeiras peitorais das raias e são eletro-receptores sensíveis. O canal que conecta a célula receptora e o poro é preenchido por um gel condutor de eletricidade enquanto que a parede do canal não é condutora.
Como o canal se estende para o interior da pele a célula é capaz de perceber as alterações do campo elétrico do exterior, quando comparado com o tecido onde as células estão apoiadas. Deste modo, estas células funcionam como voltímetros que detectam alterações no campo elétrico ao redor do corpo. A sensibilidade a alteração do campo elétrico destas células é notável, alcançando a sensibilidade de 0,01 microvolt por centímetro, alteração capaz de ser detectada somente pelos mais sensíveis voltímetros industriais.
Os Elasmobranchii utilizam esta sensibilidade elétrica na detecção de presas. Um tubarão, por exemplo, tem a capacidade de detectar uma presa escondida, somente reconhecendo as alterações no campo elétrico que a presa causa. Toda atividade muscular gera um potencial elétrico, além da própria geração do campo elétrico resultado da interação química entre o animal e a água circundante.
Este mesmo sentido pode ser utilizado por este grupo para a navegação. A Terra apresenta um campo magnético, sendo que as linhas do campo gerado apresentam voltagem suficiente para ser percebida pelas ampolas de Lorenzine. Do mesmo modo as correntes oceânicas apresentam campos elétricos suficiente para a detecção pelas ampolas conforme movimentam íons dissolvidos na água.
(https://siteantigo.portaleducacao.com.br/conteudo/artigos/biologia/o-sistema-da-linha-lateral/36085)
O sistema da linha lateral é uma intrincada série de receptores mecânicos dispostos lateralmente no corpo, desde a região anterior (cabeça) até a região caudal. Estes receptores são sensíveis ao som, toque, pressão e movimento. Funões: detectar presa, evitar predador, formação de cardumes (estímulos mecânicos oriundos do meio).
A unidade básica da linha lateral é o neuromasto, que são grupamentos de células ciliadas sensíveis ao deslocamento mecânico. Na maioria dos peixes os neuromastos estão arranjados em uma série de canais na cabeça e na lateral o corpo chegando até a cauda. Este mesmo sistema é encontrado nas larvas aquáticas dos anfíbios e nos adultos dos anfíbios aquáticos, mostrando-se como uma condição plesiomórfica dos tetrápodes.
https://youtu.be/z0M7_HPSi14
Peixes eletricos
Orelha interna
Os peixes apresentam também, como todo vertebrado, uma orelha interna capaz de detectar sons, todavia, o sistema de linha lateral, em geral, é o sentido mais aguçado.
A unidade básica da linha lateral é o neuromasto, que são grupamentos de células ciliadas sensíveis ao deslocamento mecânico. Na maioria dos peixes os neuromastos estão arranjados em uma série de canais na cabeça e na lateral o corpo chegando até a cauda. Este mesmo sistema é encontrado nas larvas aquáticas dos anfíbios e nos adultos dos anfíbios aquáticos, mostrando-se como uma condição plesiomórfica dos tetrápodes.

Relação entre os Ossículos Weberianos e o ouvido interno

Os peixes são, também, um dos primeiros vertebrados a desenvolverem órgãos de audição, neste caso, um "ouvido" primitivo que compreende somente a parte "interna" do ouvido, sem as estruturas que fariam parte dos ouvidos médio e externo. Desta maneira, a audição como a percepção de "sons" em si não é uma das mais aguçadas nos peixes, porém, outro sistema que funciona a base de um princípio semelhante ao da audição e que é encontrado nos peixes, servindo como um "complemento" à sua audição pouco eficiente, é a linha lateral. A linha lateral seria uma espécie de "vinco" que se estende ao longo do comprimento do animal em suas faces laterais desde o opérculo até a cauda marcado por vários poros que levam até uma série de células ciliadas que conseguem perceber "distúrbios" na água que entra pelos poros na forma de vibrações (assim como os ossículos dos ouvidos vibram quando estimulados pelo ar em deslocamento, gerando o som), assim, podem identificar quando algo se desloca debaixo da água (um predador, uma presa...), em que ambiente se encontra, qual a sua profundidade na água, etc. Embora o ouvido dos peixes não seja muito adequado para a percepção de sons, estes também auxiliam na sensação de equilíbrio, contribuindo para uma melhor locomoção do animal. Assim como no caso do olfato, a acidificação do mar de maneira descontrolada também pode levar ao prejuízo da audição nos peixes, já que afeta o cálcio que compõe os otólitos do ouvido interno destes animais. Além disso, a poluição sonora dos ambientes urbanos afeta os peixes na medida em que "abafa" os sons que são de maior importância para eles, como o de um predador vindo em sua direção, ou de uma presa indefesa nadando; provoca maior estresse nestes animais, os confundindo e fazendo-os mais propícios a ficarem indefesos ou relutantes em tomar decisões. (texto do Al. Barcellos, 2020)

Flutuabilidade
A flutuabilidade de um objeto imerso é dado pela contraposição de duas forças opostas, empuxo e gravidade. A gravidade é função da massa do objeto, e o empuxo é função do seu volume. Desse modo, quanto menor a diferença entre a densidade total do objeto e a densidade do meio líquido em que se encontra, maior sua flutuabilidade, e de modo oposto, quanto maior a diferença entre sua densidade total e a densidade do meio líquido em que se encontra, menor sua flutuabilidade.
Os mecanismos de flutuabilidade em peixes envolvem a diminuição da densidade total do corpo pela incorporação de componentes de baixa densidade (gases, óleos) em diferentes órgãos.
órgão hidrostático chamando bexiga natatória, cuja função principal é auxiliar na flutuabilidade do peixe, através do controle do peso específico corporal do peixe em relação ao peso da água. Assim, há um aumento de volume quando o peixe sobe na coluna d'água, e quando o peixe desce, uma diminuição de volume.
A bexiga natatória é um divertículo faríngeo preenchido por gases, que nos Neopterygii tem origem da porção dorsal da faringe; ela pode ou não manter ligação com a faringe; quando a ligação entre a bexiga natatória e a faringe é mantida, tem-se a condição fisóstoma, presente nos Holostei e Teleostei mais basais; Quando essa condição é perdida nos adultos, tem-se os fisóclistos.
No primeiro caso, a bexiga natatória é preenchida por ar engolido pela boca, enquanto no segundo caso, a bexiga natatória é preenchida por gases secretados a partir do sangue.
Os fisóclistos desenvolveram mecanismos especiais de difusão de gases entre a bexiga natatória e o sangue, baseados em áreas altamente vascularizadas na parede da bexiga (rede mirabile): processos bioquímicos levam à difusão de gases como CO2, O2, H2 e N2 do sangue para dentro da bexiga natatória, principalmente num ponto chamado glândula de gás, o que aumenta a flutuabilidade; a composição de gases na bexiga natatória é diferente da do ar, variando entre os diferentes grupos de peixes. Outra região da parede da bexiga natatória, igualmente vascularizada, conhecida por glândula oval, é responsável pelo movimento inverso de gases, desinflando a bexiga quando o peixe quer aprofundar-se.
Os tubarões, ao contrário dos osteíctes, que podem regular através da bexiga natatória sua densidade, não apresentam uma bexiga natatória, que podem encher de gás diminuindo sua densidade ou aumentando-a. Usando a analogia de um dirigível ou zepelim, um peixe ósseo pode ser comparado a um dirigível, enquanto um tubarão é mais parecido com um avião. Os tubarões dependem da sustentação gerada por suas grandes nadadeiras dorsais da mesma forma que as asas de um avião fornecem sustentação ao voo, no entanto, essa sustentação não seria suficiente por si só para sustentar o tubarão com o tamanho de nadadeiras que eles têm (a menos que se movam rápido), muito da elevação (menor profundidade, ou subir em direção a superfície, ou dentro d'água, dos tubarões vem de seu fígado. Os tubarões têm fígados enormes, estes contêm óleo que é menos denso que a água e, portanto, flutua, o que os tornam ligeiramente negativamente flutuantes. Ao contrário da crença popular, os tubarões não precisam nadar constantemente ou morrem; no entanto, eles precisam nadar para evitar afundar.
O fígado cheio de óleo, ao contrário de uma bexiga natatória, significa que os corpos dos tubarões são incompressíveis, permitindo que eles se movam entre águas muito profundas e a superfície com facilidade. Esses peixes ósseos que vivem em altas pressões geralmente morrem quando trazidos à superfície, os tubarões não. Pouco se sabe sobre os tubarões de águas profundas, mas o que se sabe é que eles são abundantes, a fisiologia básica do tubarão é bem adequada para a vida tanto em profundidade quando mais à superfície da água.
OSTEICHTHYES
Peixes ósseos

Original National Museum of Natural History exhibit of a coelacanth, Latimeria chalumnae, that went on display on August 20, 1957. This was a model of a male fish caught off the Comoro Islands, near Madagascar.

Celacanto fóssil

Fóssil completo de celacanto (peixe ósseo), do tamanho de um tubarão branco, o maior ja encontrado, que viveu ao período Jurássico
(Universidade de Portsmouth. exame).

Celacanto (domestika)

Marjorie Courtenay-Latimer com sua descoberta em 1938.
(WP)

Nature, março de 1939

Latimeria spp

Marjorie Courtenay-Latimer included this sketch in a letter sent to J.L.B. Smith, requesting his help in identifying an unusual fish brought in as by-catch on a local fishing vessel. Note that Courtenay-Latimer drew the tail (labeled “caudal fin” here) twice to bring attention to that distinctive feature. Image credit: African Coelacanth Ecosystem Programme (ACEP)/ South African Institute for Aquatic Biodiversity (SAIAB).(Fonte: Curious Sengui)

a) Mawsonia, b) L. chalumnae, c) L. menadoensis

The coelacanth propels itself forward using its second dorsal and first anal fins, as marked by the arrows. These two fins beat in unison. This mode of swimming is very similar to that found in [the completely unrelated] Ocean Sunfish (Mola mola). The paired pectoral and pelvic fins serve more to stabilize the animal. Image credit: YouTube. (1)








In 1938, Marjorie Courtenay-Latimer, the first curator of the Natural History Museum in East London, South Africa, was presented a breathtaking, well-preserved coelacanth (pronounced SEAL-a-canth) by Captain Hendrik Goosen.
Coelacanths were presumed to have died off en masse like the dinosaurs during the last great extinction at the end of the Cretaceous period some 63 million years ago.
From the moment Professor J.L.B. Smith, Rhodes University, South Africa, classified this rare living fossil Latimeria (after the East London curator) chalumanae (from the Chalumna River were it was thought to be found) a modern day mystery began to slowly unravel snippets of an ancient creature that seems to have trumped evolution.
In 1839, Louis Agassiz found the first fossil of a coelacanth in a Permian (290-240 million years ago) marl slate in Durham, northern England. He discovered that the fin rays supporting the tail were hollow and named it Coelacanthus (from the Greek for hollow spine) granulatus (after the tubercular ornamentation of its scales).
Other fossils around the globe depicted an extraordinary diversity of Devonian (410-360 million years ago) species that were as large as 10 feet and recent Cretaceous (138-63 million years ago) specimens were as small as one foot.
Prior to 410 million years ago, only a few spiky, low plants, along with some scorpions and other insects, lived on land. The continents were unrecognizable: large fresh water lakes and the oceans brimmed with life.
The diversity of aquatic life was dazzling, including: small, flat, heavily armored, bottom-feeding jawless bone-skinned fishes; giant nautilus, the size of a person; sea scorpions larger than lobsters; the first jawed fish; and primitive giant sharks - even then fearsome predators.
The fish of 400 million years ago were separated into two classes: ray finned with single dorsal fins and paired pectoral and pelvic fins common in modern fishes; and the lobed finned fishes - the coelacanth, the lungfish and the rhipidistian whose fins seemed to grow from the end of fleshy, limb-like lobes or toeless legs. Many were predators all were abundant.
About 360 million years ago, a fresh water lobed-finned fish sprouted legs, crawled out of the water and conquered the land. Since the time of Louis Agassiz and Charles Darwin scientists have been trying to discern which group: lungfish, rhipiditian or coelacanth, evolved legs.
Due to WW II, Professor Smith was forced to wait to get his next sample. He went to great lengths to distribute reward pamphlets all along the East African coast. Meanwhile Comoran fishermen had been occasionally catching coelacanths unaware of their scientific value. So it was only the next known to science that was caught in 1952 off the Comoro Islands, a remote volcanic archipelago at the head of the Mozambique Channel, halfway between Mozambique and Madagascar.
The anatomy of these primeval creatures is truly worthy of admiration and has allowed scientists a rare glimpse through the looking glass of 400 million years of time. There are about 26,000 species of living fish and easily the coelacanth is the most mysterious.
The paired pectoral and pelvic fins appear similar to those of a lizard.
Coelacanths are able to swim forward, backward, upside down and on their heads.
They have an extremely small brain (1/15,000 of an adults weight) with a unique intracranial joint, a feature previously known only from fossils. The coelacanth is able to increase its gape by a sudden opening of the entire head, giving it a powerful bite and compensating for its comparatively small, sharp teeth.
A rostal organ is jelly-filled with three exterior openings on either side of its snout - unknown in any other living creature. It acts as an electro-detector constantly searching for prey.
Coelacanths generally live below 1,300 feet, although they have been observed swimming as closes as 100 feet from the surface. During the day, they are inactive, seeking protection in volcanic caves. At night, these nocturnal predators are on the hunt. Their luminescent alien-green eyes have adapted to extremely low light levels.
Their spines are filled with a low viscosity lipid and the coelacanth is composed mainly of a flexible cartilage - similar to a shark. Its spinal fluid is viscous golden and rumored to act as an elixir. Coelacanths are thought to live for at least 40 years. The coelacanth's swim bladder is a slender fat-filled tube embedded in fat below the spine - it serves to increase its buoyancy. Buoyancy is maintained by the entire lipid-filled body of the fish and so, like many sharks, this ancient fish is capable of considerable vertical movement in the water column.
Coelacanths give birth to live pups, this predates the first mammals by at least 100 million years.
In 1987, a submersible located and filmed wild coelacanths off the Comoro Islands. Since then, coelacanths have been filmed or photographed in their habitat by divers and ROVs in Indonesia, Madagascar, South Africa and along the Tanzanian coast.
The Convention on International Trade in Endangered Species (CITES) listed the coelacanth in 1989 as severely endangered and strictly forbids any trade in this animal. In 1998, Professor Mark Erdman, University of California Berkeley, discovered a new golden brown Indonesian coelacanth Latimeria menadoensis some 6,000 miles from the Comoro Islands in the Indian Ocean. In fact, the Indonesian coelacanth has the same coloration of the Indian Ocean ones: a clock spring steely blue with white to off white or pink blotches, the pattern of which is characteristic of the individual (rather like a fingerprint). Other species of coelacanths are surely awaiting discovery; reiterating a heightened need for protecting all oceans, seas, lakes and rivers.(The Malibu Times).
PEIXES ÓSSEOS
Características gerais dos peixes ósseos
Peixe ósseo, é qualquer membro da superclasse Osteichthyes, um grupo formado pelas classes Sarcopterygii (peixes de nadadeiras lobadas) e Actinopterygii (peixes com nadadeiras raiadas) do subfilo Craniata (Vertebrata), incluindo a grande maioria dos peixes vivos e praticamente todos os pescado do mundo e peixes comerciais.
O termo científico Pisces também foi usado para identificar esse grupo de peixes.
Os Osteichthyes são caracterizados por osso endocondral ("esponjoso") no endoesqueleto, raios ósseos nas nadadeira formados por lepidotriquias (escamas modificadas em forma de ladrilho) e três pares de ossos dérmicos com dentes que revestem as mandíbulas (dentário, pré-maxilar e maxilar).




Osteichthyes exclui os peixes sem mandíbula da classe Agnatha (peixes-bruxa e lampreias) e os peixes cartilaginosos que constituem a classe Chondrichthyes (tubarões, tubarões serra, raias e quimeras), mas inclui as 20.000 espécies e mais de 400 famílias de peixes ósseos modernos (infraclasse Teleostei), bem como algumas formas ancestrais.
A principal característica dos peixes ósseos é seu esqueleto, pelo menos parcialmente composto de osso verdadeiro (em oposição à cartilagem).
Outras características incluem, na maioria das formas, a presença de uma bexiga natatória (um saco cheio de ar para dar flutuabilidade), coberturas branquiais sobre a câmara branquial, escamas ósseas semelhantes a plaquetas, (ganóides, ciclóides e ctnenóides), um crânio com suturas e fertilização externa de ovos.

Crâneo de Amia calva um peixe ósseo da ordem Amiiformes .
Os peixes ósseos ocorrem em todos os ambientes de água doce e oceânicos, incluindo cavernas, habitats de águas profundas e fontes termais e respiradouros. A variedade de formas e hábitos comportamentais é notável.
Seus tamanhos de corpo variam de espécies minúsculas, como o góbio pigmeu (Pandaka pygmaea; 12,0 mm) aos enormes marlins e peixes-espada (família Istiophoridae) com comprimentos de até 4,5 m e o peixe-lua do oceano (Mola mola), que pode pesar mais de 900 kg (1 tonelada).

Sun fish ou peixe-lua Mola mola
(Fonte: Ilse Reijs and Jan-Noud Hutten)

Reconhecem-se atualmente três espécies válidas, Mola mola (Linnaeus, 1758), Mola alexandrini (Ranzani 1839) e Mola tecta (Nyegaard, 2017). Com a revisão de literatura sobre o gênero Mola, descobriu-se que as espécies Mola alexandrini e Mola ramsayi (Gigliogi, 1883) são sinônimos, mas como a descrição M. ramsayi foi posterior, o nome que deve ser usado é Mola alexandrini respeitando-se o principio da prioridade em taxonomia.
(Fonte imagem: Emma Burns)

Características gerais
dos peixes ósseos
Nadadeiras com lepidotríquias (raios);
Presença de pulmão ou bexiga natatória derivados do tubo digestivo;
Padrão de ossificação da cintura peitoral;
Modificações no padrão da musculatura da faringe e da mandíbula.
Sinapomorfias dos sarcopoterigi
Junta intercraniana;
Nadadeiras pares musculares do tipo arquipterígio;
Dentes recobertos por esmalte.
Clado Sarcopterigii
Dipnoi (peixes pulmonados)
Caracterísiticas
Narinas de posição interna.
Respirção pulmonar e branquial.
Placas dentarias.
Escamas cosmoides adelgazadas en formas actuales.
Notocorda persistente en formas actuales.
Aleta arquipterigia
Ovíparos, depredadores.
Agua dulce, África, Australia y Sudamérica.
Nadadeira dorsal única (originalmente; pode estar dividida em alguns Teleostei);
Elementos basais da nadadeira peitoral aumentados;
Dentes recobertos na extremidade por acrodina (tecido originário tanto do ectoderma como de células do ectomesêmquima) e na base por esmalte (derivado do ectoderma);
Raios das nadadeira ímpares articulam-se com elementos esqueléticos internos ao corpo, sem a interveniência de lobos basais;
Escamas com padrão típico de articulação e revestinas por ganoína (presente nos grupos mais basais);
Perda na cistura pélvica e sua substituição por parte do metapterígio;
Telencéfalo invertido;
Seqüências de nucleotídeos características.

















Elementos basais da nadadeira peitoral aumentados;
Dentes recobertos na extremidade por acrodina (tecido originário tanto do ectoderma como de células do ectomesêmquima) e na base por esmalte (derivado do ectoderma);
Raios das nadadeira ímpares articulam-se com elementos esqueléticos internos ao corpo, sem a interveniência de lobos basais;
Escamas com padrão típico de articulação e revestinas por ganoína (presente nos grupos mais basais);
Perda na cistura pélvica e sua substituição por parte do metapterígio;
Telencéfalo invertido;
Seqüências de nucleotídeos características.
Forma hidrodinâmica
Corpo fusiforme, i.e., uma forma corporal que apresenta as extremidades mais estreitas que o centro. Esse formato, dito hidrodinâmico promove a redução do atrito com a água, proporcionando um deslocamento com menos gasto de energia pois com menos arraste (atrito) com o ambiente.


Forma hidrodinâmica
Muco e escamas
Escamas ganóides, ciclóides e ctenóides
O muco, secretado por glândulas mucosas tem duas funções básicas: facilitar o deslocamento na água e proteger o corpo da entrada de bacterias e agentes patogênicos.
As escamas atuam para melhorar o deslocamento na água, e por serem impermeáveis dificultam a saída de sais do corpo do peixe (de água doce), atuam como proteção contra predadores e patógenos.


Tipos de escamas dos peixes: escamas ganóides de pirarucu (Arapaima gigas)*, escamas cosmóides de peixe-lagarto (Lepisosteus oculatus)**, e escamas ósseas do tipo ciclóides de voga (Cyphocarax voga) e ctenóides de peixe-rato (Caelorinchus marinii). (Fonte:* em domínio público, autor: George Chernilevsky; ** autor: Stan Shebs (ambas de Wikimedia.org)).

Exemplos de revestimentos nos peixes: (a) somente por pele em bagre (Genidens genidens); (b) escamas ganóides em esturjão (Scaphyrhynchus suttkusi*), (c) escamas ciclóides em voga (Cyphocarax voga); (d) escamas ctenóides em peixe-rato (Caelorinchus marinii); (e) escudos laterais em xaréu (Caranx hippos); (f) escamas em forma de escudos no voador (Dactylopterus volitans); (g) escudos ventrais em Gephyroberyx darwinii; (h) placas ósseas na cascuda (Hoplosternum littorale); (i) placas retilíneas de escamas no peixe-porco (Balistes capriscus);
(j) tubérculos ósseos no peixe-morcego (Ocgocephalus vespertilio); (k) espinhos ósseos no baiacu-de-espinhos (Cyclichthys spinosus); (l) séries de anéis ósseos formados por placas ósseas no peixe-cachimbo (Syngnathus folleti). * foto do esturjão de Patrick O'Neil, Geological Survey of Alabama, e detalhe das escamas de NOAA Fisheries Service, ambas em Domínio Público. (Fonte: Fishcer, L.C.)
Boca anterior (terminal) maioria.


(Blog da ex-aluna Alessandra Dorigon)
Intestino longo com cecos pilóricos.
Bexiga natatória
Hidrostase: Equilíbrio dependente da densidade da água.
Responsável pela flutuação do peixe, permitindo que ele suba ou desça na coluna d'água sem usar os músculos.
Recepção e percepção de sons (órgão de Webber).
Percepção da pressão dependente da profundidade.
Respiração: fisóstomos suplemento na respiração branquial.
Dipnóicos, e também em períodos de seca onde habitam, em lodo.





Bexiga natatória (swim bladder, By Jon Houseman)
A vesícula gasosa possui uma glândula que permite a liberação e a introdução de gases, principalmente oxigénio, neste órgão, para aumentar o seu volume.
O mecanismo para a troca de gases é o seguinte: a glândula excreta ácido láctico que provoca a liberação ou a perda do oxigénio da hemoglobina.
Este gás difunde-se para dentro da vesícula gasosa através de uma complexa estrutura chamada "rete mirabile". Em outra região da vesícula, existe um órgão chamado "janela oval". Nesse órgão o gás entra em contato com o sangue. É através da janela oval que o oxigênio pode voltar para a corrente sanguínea, diminuindo desta forma a pressão dentro da bexiga natatória e diminuindo o seu volume e consequentemente aumentando a densidade do peixe. A proporção de gases dentro da vesícula gasosa varia (ou bexiga natatória): em águas pouco profundas, a composição é muito próxima da da atmosfera, enquanto que em águas profundas tem a tendência para ter uma maior percentagem de oxigênio. Observou-se, por exemplo, na enguia Synaphobranchus (que já foi encontrada a uma profundidade de 5440 metros) a seguinte composição do gás da sua bexiga natatória: 75,1% de oxigênio, 20,5% de nitrogênio, 3,1% de dióxido de carbono e 0,4% de argônio.

Localização da bexiga nataória (Fonte E. Brit.)
A bexiga natatória normalmente consiste em dois sacos cheios de gás localizados na porção dorsal do peixe, embora em algumas espécies primitivas haja apenas um único saco. Possui paredes flexíveis que se contraem ou se expandem de acordo com a pressão ambiente. As paredes da bexiga contêm poucos vasos sanguíneos e são revestidas por cristais de guanina, que as tornam impermeáveis aos gases. Ao ajustar o órgão de pressurização de gás usando a glândula de gás ou janela oval, os peixes podem obter flutuabilidade neutra e subir e descer a uma grande variedade de profundidades.
Devido à sua localização próxima a posição dorsal confere estabilidade lateral ao peixe. Nas bexigas natatórias fisostômicas (peixes fisóstomos), uma conexão é mantida entre a bexiga natatória e o sistema digestório, o duto pneumático, permitindo que os peixes encham a bexiga natatória "engolindo" o ar atmosférico. O excesso de gás pode ser removido de maneira semelhante. Em variedades mais derivadas de peixes (os peixes fisóclistos), a conexão com o trato digestivo é perdida. Nos primeiros estágios da vida, esses peixes precisam subir à superfície para encher suas bexigas natatórias; nos estágios posteriores, o ducto pneumático desaparece e a glândula de gás tem que introduzir gás (geralmente oxigênio) retirado da corrente sanguínea, na bexiga para aumentar seu volume e, assim, aumentar a flutuabilidade. Para introduzir o gás na bexiga, a glândula gasosa excreta ácido lático e produz dióxido de carbono. A acidez resultante faz com que a hemoglobina do sangue perca seu oxigênio (efeito Root), que então se difunde parcialmente para a bexiga natatória. O sangue que flui de volta para o corpo entra primeiro em uma órgão chamado rete mirabile, onde praticamente todo o excesso de dióxido de carbono e oxigênio produzidos na glândula de gás se difunde de volta para as artérias que fornecem a glândula de gás. Assim, uma pressão de gás de oxigênio muito alta pode ser obtida, o que pode até ser responsável pela presença de gás nas bexigas natatórias de peixes de águas profundas como a enguia, exigindo uma pressão de centenas de bares. Em outra região da bexiga, em uma estrutura semelhante chamada de janela oval, o gás da bexiga está em contato com o sangue e o oxigênio pode se difundir novamente para a corrente sanguínea. Junto com o oxigênio, outros gases são eliminados na bexiga natatória, o que também é responsável pelas altas pressões de outros gases. A combinação de gases na bexiga varia entre as espécies.
Em peixes de águas rasas, as proporções se aproximam da atmosfera, enquanto peixes de águas profundas tendem a ter porcentagens mais altas de oxigênio. Por exemplo, foi observado que a enguia Synaphobranchus sp. tem 75,1% de oxigênio, 20,5% de nitrogênio, 3,1% de dióxido de carbono e 0,4% de argônio em sua bexiga natatória. As bexigas natatórias fisóclistas têm uma desvantagem importante: proíbem o aumento rápido, pois a bexiga estouraria. Os peixes fisóstomos podem "arrotar" o gás, embora isso complique o processo de re-submersão. A bexiga natatória em algumas espécies, principalmente peixes de água doce (carpa comum, bagre, etc.) está interconectada com o ouvido interno do peixe. Eles são conectados por quatro ossos chamados ossículos weberianos do aparelho weberiano. Esses ossos podem transportar as vibrações para o sáculo e a lagena. Eles são adequados para detectar sons e vibrações devido à sua baixa densidade em comparação com a densidade dos tecidos do corpo do peixe. Isso aumenta a capacidade de detecção de som. A bexiga natatória pode irradiar a pressão do som, o que ajuda a aumentar sua sensibilidade e expandir sua audição. Em alguns peixes de águas profundas, como o Antimora sp, a bexiga natatória também pode estar conectada à mácula do sáculo para que o ouvido interno receba uma sensação da pressão do som. Na piranha de barriga vermelha, a bexiga natatória pode desempenhar um papel importante na produção de som como ressonador. Os sons produzidos pelas piranhas são gerados por contrações rápidas dos músculos sônicos e estão associados à bexiga natatória.(Taylor et all, (2010) e em Wikipedia Taylor et all, 2010).

Swim bladders: (a) a physostomous fish with swim bladder connected to the digestive system by the pneumatic duct; (b) vascular connections of a physoclistous swim bladder which is disconnected from the digestive system;
(c) the physoclistous swim bladder of Seriola hippos which is connected externally by the swim bladder vent. N.B. (a) and (b) from Pough et al. 1996.
A flutuabilidade de um objeto ou ser vivo imerso em água é dado pela contraposição de duas forças opostas: a gravidade e o empuxo.
A gravidade é função da massa do objeto, e o empuxo é função do seu volume. Um objeto que flutua em um líquido é positivamente flutuante. Isso significa que a quantidade de água deslocada pelo objeto pesa mais do que o próprio objeto. Assim, quanto menor a diferença entre a densidade total do objeto e a densidade do meio líquido em que se encontra, maior sua flutuabilidade. Um objeto cujo peso é superior ao peso do líquido que ele desloca, não poderá flutuar, nesse caso afundará. Quanto maior a diferença entre sua densidade total e a densidade do meio líquido em que se encontra, menor sua flutuabilidade.
Os mecanismos de flutuabilidade em peixes envolvem a diminuição da densidade total do corpo pela incorporação de componentes de baixa densidade (gases, óleos) em diferentes órgãos.
Sistema excretor e Excreção
Peixes ósseos apresentam rins mesonéfricos e excretam amônia
Experimento para fazer em casa e trazer os resultados para aula:
Leia o texto desse artigo e construa o modelo, faça os experimentos e anote os resultados. Floating with swim bladder. (Svenja Lohner, 2018).
Bexiga natatória, órgão de equilíbrio hidrostático (que pode ser de dois tipos: fisóstoma e fisóclista).
A bexiga natatória dos peixes teleósteos é embriologicamente derivada do trato digestivo e em alguns peixes é mantida uma comunicação com ele através de um ducto pneumático graças ao qual é regulado o volume do gás no interior do órgão (bexigas natatórias fisóstomas (do grego, physa = bexiga, stoma = boca).
Em muitos peixes teleósteos esta comunicação se perdeu e a bexiga natatória passou a ser um órgão fechado, (bexiga fisóclista onde o gás é secretado por uma glândula constituída de um epitélio secretor associado a uma rede capilar muito desenvolvida chamada de rete mirabile. Muitos fisóstomos também possuem o sistema glandular rete mirabile para a secreção de gás. (Jones & Marshall, 1953).
A bexiga natatória é um divertículo faríngeo preenchido por gases, que nos Neopterygii tem origem da porção dorsal da faringe; ela pode ou não manter ligação com a faringe; quando a ligação entre a bexiga natatória e a faringe é mantida, tem-se a condição fisóstoma, presente nos Holostei e Teleostei mais basais; Quando essa condição é perdida nos adultos, tem-se os fisóclistos. No primeiro caso, a bexiga natatória é preenchida por ar engolido pela boca, enquanto no segundo caso, a bexiga natatória é preenchida por gases secretados a partir do sangue.
Os fisóclistos desenvolveram mecanismos especiais de difusão de gases entre a bexiga natatória e o sangue, baseados em áreas altamente vascularizadas na parede da bexiga (rede mirabile): processos bioquímicos levam à difusão de gases como CO2, O2, H2 e N2 do sangue para dentro da bexiga natatória, principalmente num ponto chamado glândula de gás, o que aumenta a flutuabilidade; a composição de gases na bexiga natatória é diferente da do ar, variando entre os diferentes grupos de peixes.
Outra região da parede da bexiga natatória, igualmente vascularizada, conhecida por glândula oval, é responsável pelo movimento inverso de gases, desinflando a bexiga quando o peixe quer aprofundar-se.
Peixes com bexiga natatória podem encher a bexiga respirando o ar na superfície. Entretanto, peixes que vivem a grandes profundidades isso é impraticável e impossível, pois um grande volume de ar teria que ser acumulado dentro da bexiga na superfície para atingir a flutuabilidade neutra em grandes profundidades, onde a pressão é muitas vezes maior.
Bexiga natatória superinsuflada ou rompida:
Instabilidade. Incapacidade dos peixes de submergir, olhos esbugalhados Protrusão de órgãos internos, que aparecem pela boca. Sangramento interno e externo e possível morte.
Órgãos dos sentidos
Olhos e linha lateral.
Nadadeiras pares
Um par de nadadeiras peitorais
Um par de nadadeiras pélvicas.
Nadadeiras impares
Dorsal, anal e caudal.
Reprodução
Apresentam sexos separados, e cada indivíduo possui gônadas geralmente pares. A grande maioria é ovípara com fecundação externa, embora existam espécies com fecundação interna e hermafroditas.
Algumas espécies passam por mudanças de sexo, com machos que passam a fêmeas aumentando de tamanho e as fêmeas que se tornam dominantes nos cardumes, ao passarem a machos. Os ovos são pequenos e sem anexos embrionários mas com quantidade de vitelo variável entre as espécies.
Desenvolvimento indireto: estágio larvar: alevino

Alevino de tambaqui Colossoma macropomum.
Fonte: Desenvolvimento embrionário de tambaqui (Lima, 2014)
Ovulíparos
Apresentam saco vitelínico.
Tegumento com escamas e glândulas mucosas
Muco: diminuição do atrito do corpo com o meio e proteção contra micro-organismos.
Tipos de escamas: escamas ctenóides, ciclóides, ganóides, cosmóides
Sistema digestório
Sistema digestório completo e digestão extracelular, intestino em forma de S e sem cloaca.
Diversidade nos hábitos alimentares:
filtradores, frugívoros, herbívoros, insetívoros, carnívoros.
Sistema respiratório e trocas gasosas:
Quatro pares de brânquias
Opérculo protegendo as brânquias (Opérculo é uma placa óssea que recobre as brânquias).
Respiração pulmonar
Peixes dipnóicos: nos peixes ósseos dipnóicos (ex: pirambóia) a bexiga natatória atua como um pulmão.
Sistema circulatório
A circulação é fechada e simples,
Hemácias ovais e nucleadas
Coração com duas cavidades: uma aurícula e um ventrículo
No coração dos peixes só circula sangue venoso (vindo do corpo para as brânquias) e depois de oxigenado (i.e., depois da hematose) o sangue segue para todo o copro do animal.
Crânio com suturas cranianas
Nas suturas as extremidades dos ossos têm inter-digitações ou sulcos, que os mantêm íntima e firmemente unidos. Conseqüentemente, as fibras de conexão são muito curtas preenchendo uma pequena fenda entre os ossos. Este tipo de articulação é encontrado somente entre os ossos planos do crânio. Na maturidade, as fibras da sutura começam a ser substituídas completamente, de ambos os lados da sutura tornam-se firmemente unidos/fundidos. Esta condição é chamada de sinostose.
Função da brânquia na osmorregulação
Problema Básico:
O processo de osmose faz com que o sangue dos peixes de água doce tenha uma pressão osmótica mais alta do que a água em que nadam.
A pressão osmótica é expressa em miliosmoles. O sangue de um peixe de água doce tem aproximadamente 300 mOsmol / litro de água, enquanto a água doce do meio onde vivem geralmente tem menos de 5 mOsmol / l.

Portanto, os teleósteos dulcícola são hiperosmóticos em relação a seu ambiente, ou seja, seu fluido corporal (principalmente seu sangue) tem um conteúdo de soluto mais alto que o ambiente (o sangue é mais concentrado em sais do que a água do ambiente circundante). O resultado é uma forte tendência da água se difundir no peixe de água doce (a água tende a entrar no corpo) e os sais tendem a sairem do corpo.
Os peixes podem resistir a esse movimento osmótico por terem uma cobertura corporal relativamente impermeável, pele e escamas ajudam nesse aspecto, entretanto, a membrana epitelial das brânquias deve ser altamente permeável para que as trocas gasosas ocorram. Assim, a água se difunde entrando pela membrana epitelial das brânquias e os peixes tendem a se hidratar.

Com os peixes de água salgada, o problema é inverso. Os peixes de água salgada são hiposmóticos em relação à água do mar, seu sangue tem um conteúdo de soluto menor e, portanto, uma pressão osmótica mais baixa (cerca de 400 mOsmol) do que a água do mar (que tem cerca de 1000 mOsmol).
Os peixes marinhos sofrem uma perda passiva de água pelas brânquias e um ganho passivo de sais. Peixes de água salgada tendem a desidratar.
Para manter os 300 mOsmol /L em seu sangue, apesar da tendência osmótica de ganhar água e perder íons, um peixe de água doce deve procurar captar ativamente íons do ambiente e excretar água de seu corpo. Ele consegue isso:
1) interrompendo a saída de sal pelas brânquias e
2) produzindo uma urina abundante e diluída.
Os peixes de água doce nunca bebem, pois ganham toda água de que precisam (e muito mais) passivamente pelas brânquias que necessitam ser altamente permeáveis para que possa trocar o CO2 do sangue pelo O2 diluído na água.
A teoria aceita de como os peixes dulcícolas invertem o fluxo de sal nas guelras evoluiu com o tempo. Pesquisas iniciais indicaram que enzimas (ATPases) no tecido branquial quebram o ATP e movem os sais contra o gradiente osmótico com a energia liberada.
Presumia-se que a maior parte dessa atividade ocorria em células especializadas do epitélio branquial, chamadas células de cloreto beta (para diferenciá-las das células alfa cloreto anatomicamente diferentes encontradas em peixes de água salgada). As células de cloreto têm muitas mitocôndrias, capazes de produzir uma grande quantidade de energia potencial na forma de ATP que pode ser utilizada pelas ATPases.
Esta teoria era intuitivamente atraente porque as ATPases podiam trocar Na+ por NH4+ e Cl- por HCO3-. A troca de cargas semelhantes promoveria o equilíbrio ácido-base e ambos NH4+ e HCO3- são , subprodutos residuais modificados do metabolismo. As células de cloreto, como as células epiteliais (ver cap. IIF), contêm a enzima anidrase carbônica. Parecia lógico que o dióxido de carbono e a amônia se difundissem do sangue para a célula de cloreto e o dióxido de carbono, catalisado pela anidrase carbônica, se dissociasse em bicarbonato e hidrônio. O hidrônio ionizaria a amônia e o amônio (NH4 +) e o bicarbonato (HCO3-) seriam excretados por Na+ -NH4+ ATPase e Cl- HCO3-ATPase conforme o sal era bombeado para dentro. Além disso, a Na+ -K+ ATPase localizada na membrana interna ajudaria a mover o Na+ do citoplasma para o sangue.
Pesquisas adicionais, entretanto, indicaram que outras enzimas e um gradiente eletroquímico também estavam envolvidos. Uma teoria mais recente sustenta que, embora todos os movimentos descritos acima ocorram, sem dúvida, o Na+ também segue um gradiente elétrico (a célula sendo carregada mais negativamente do que a água) para dentro da célula. O efeito combinado do gradiente elétrico e do gradiente osmótico oposto é denominado gradiente eletroquímico, e é próximo a 0 para peixes de água doce, interrompendo o fluxo de sal nas brânquias. Este potencial elétrico é mantido por uma ATPase diferente que bombeia H+ para fora da célula na água e Na+ -K+ ATPase que troca 3 Na+ por 2 K+ na membrana interna. Não está claro quais desses movimentos podem ocorrer na célula de cloreto e quais podem estar localizados nas células epiteliais.
O CO2 se difunde do sangue para a célula de cloreto, onde reage para formar HCO3-. Na verdade, a maior parte do dióxido de carbono já está na forma de bicarbonato quando atinge as brânquias, porque no pH do sangue, o HCO3- é o íon predominante e a reação de CO2 para HCO3- é catalisada pela enzima anidrase carbônica nas células vermelhas do sangue. A célula de cloreto também contém anidrase carbônica para catalisar qualquer CO2 remanescente em HCO3- e é isso que é mostrado. Obviamente, o HCO3- que já está no sangue simplesmente se difunde diretamente para a bomba de íons e é trocado por Cl-. Como não há anidrase carbônica na água, a lenta conversão de volta para CO2 não ocorre até que a água tenha saído brânquia. Portanto, o CO2 não se difunde de volta para os peixes.
Para manter 400 mOsmol, apesar de um ganho passivo de sais e perda de água, os peixes marinhos devem:
1) interromper o influxo (entrada) de sal e secretá-lo ativamente na guelra e
2) beber água do mar e se hidratar com ela.
Os peixes marinhos excretam muito pouca urina, De fato alguns peixes de água salgada nem têm glomérulos nos rins.
Enquanto os peixes de água doce e água salgada enfrentam gradientes osmóticos semelhantes (300-400 mOsmol) entre seu sangue e o ambiente, a atividade de ATPase nas brânquias do peixe água salgada é relativamente maior. A razão para isso é que o sal deve ser bombeado ativamente para o peixe no intestino para que a hidratação ocorra. Assim, nas brânquias, os peixes água salgada devem livrar seu corpo do sal adquirido ativa e passivamente. Além disso, pode ser necessário para os peixes água salgada empregar bombeamento de ATPase de Na+ em troca de NH4+ para se livrar de resíduos nitrogenados, às vezes. Obviamente, Na+ -NH4+ ATPase e Cl- HCO3-ATPase não vão ajudar na osmorregulação em peixes marinhos como se presume que façam em peixes de água doce.
Acredita-se que gradientes elétricos nos tecidos branquiais de um peixe de água doce movam os íons necessários. Nesse gradiente, a água salgada tem carga negativa, a célula alfa de cloreto é mais negativa e o sangue tem carga positiva. O gradiente elétrico é mantido pela Na+ - K+ ATPase localizada em um extenso sistema de microtúbulos. O bombeamento ativo de Na+ para fora da célula de cloreto resulta em um gradiente de Na+ (mais alto no sangue do que na célula de cloreto) que impulsiona um sistema transportador ligado a Na+ - Cl, aumentando o conteúdo de Cl- da célula e, portanto, a eletronegatividade do célula de cloreto. A célula de cloreto altamente negativa faz com que o Cl- se mova para água do mar menos carregada negativamente ao longo do gradiente elétrico. O Na+ carregado positivamente seguirá o gradiente do sangue carregado positivamente à água do mar carregada negativamente fora da célula de cloreto. Uma vez que esse mecanismo de movimentação de íons é alimentado inteiramente por Na+ - K+ ATPase, não é surpreendente que essa enzima aumente dramaticamente à medida que os peixes eurialinos passam da água doce para a salgada.
VÍDEO RESUMO SOBRE A CLASSE PEIXES
https://youtu.be/Q4yVa7YC5IM
Bibliografia
http://www.usdbiology.com/swanson/anat/Rosenfeld_Appendicular-Skeleton.pdf
Glossário importante sobre peixes:
http://australianmuseum.net.au/glossary-of-fish-terms
Origem dos peixes
http://www.infoescola.com/biologia/origem-e-evolucao-dos-peixes/
https://en.wikipedia.org/wiki/Evolution_of_fish
http://australianmuseum.net.au/glossary-of-fish-terms
Origem dos peixes
http://www.infoescola.com/biologia/origem-e-evolucao-dos-peixes/
https://en.wikipedia.org/wiki/Evolution_of_fish
http://web.unifoa.edu.br/portal/plano_aula/arquivos/04684/Aula%202%20Agnatos%20e%20Gnatostomados.pdf
Craniofacial development
https://bioone.org/journals/zoological-science/volume-22/issue-1/zsj.22.1/Craniofacial-Development-and-the-Evolution-of-the-Vertebrates--the/10.2108/zsj.22.1.full
https://bioone.org/journals/zoological-science/volume-30/issue-11/zsj.30.944/Development-of-the-Chondrocranium-in-Hagfishes-with-Special-Reference-to/10.2108/zsj.30.944.full
https://bioone.org/journals/zoological-science/volume-30/issue-11/zsj.30.944/Development-of-the-Chondrocranium-in-Hagfishes-with-Special-Reference-to/10.2108/zsj.30.944.full
Ocean Biodiversity Information System
Shark classification
Origem do Colágeno da cartilagem de Agantha
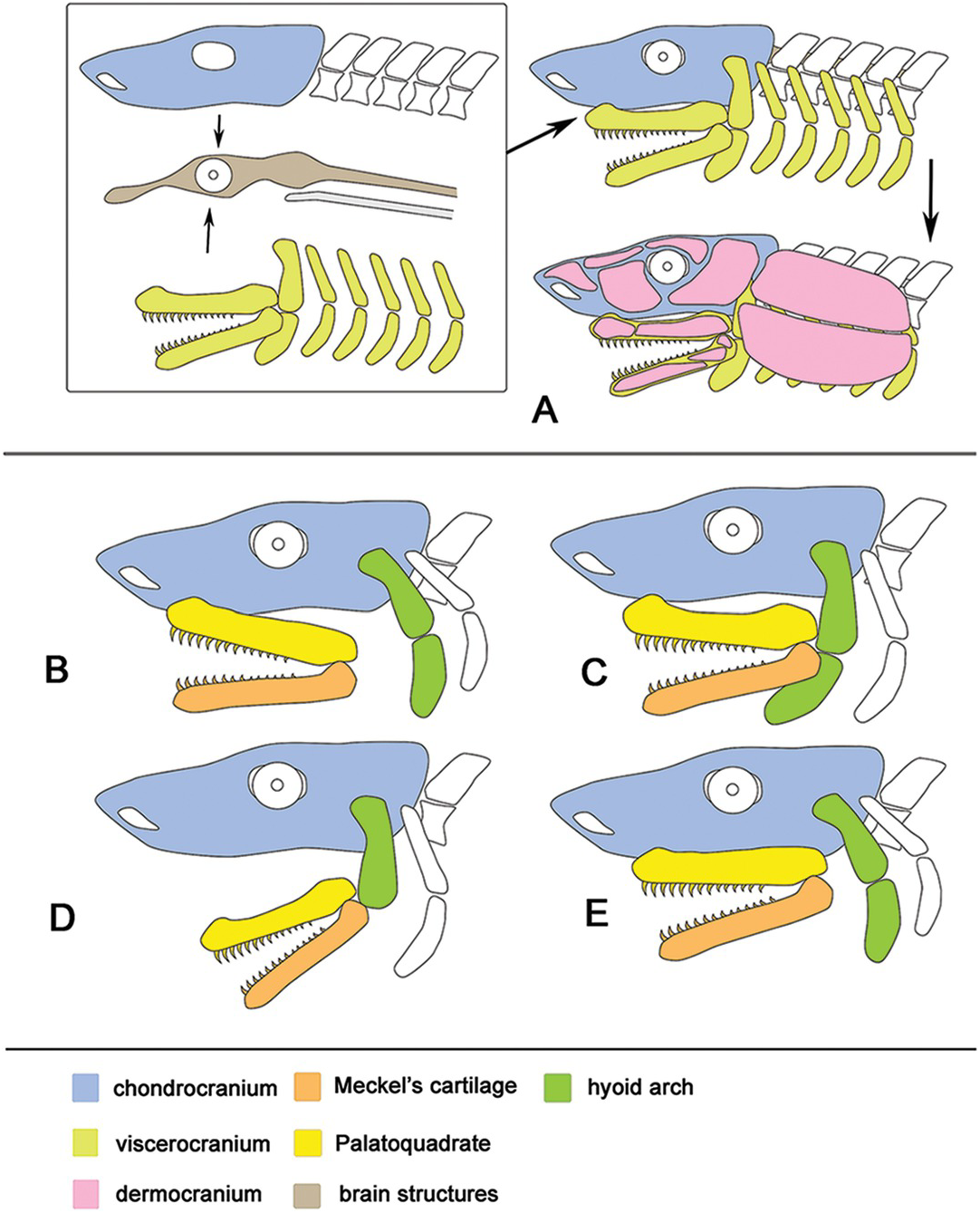{kind=link}
Sharks: biologia e diversidade
https://edisciplinas.usp.br/pluginfile.php/4414703/mod_resource/content/1/Chondrichthyes%202018_1.pdf (visitado em março de 2021)
Flutuabilidade
Trocas gasosas
Farmer, CG. 1999. The evolution of the vertebrate cardio-pulmonary system. Annual Review of Physiology 61:573-592 PDF
Farmer, CG. and D.C. Jackson. 1998. Air-breathing during activity in the fishes Amia calva and Lepisosteus oculatus. Journal of Experimental Biology 201:943-948. PDF
Farmer, C. 1997. Did lungs and the intracardiac shunt evolve to oxygenate the heart in vertebrates? Paleobiology 23(3):358-72 PDF
Equilibrio osmótico em Robalo
Coelacanth
Classificação biológica
Tráfico e contrabando
Alimentação de condríctes
Transformação das nadadeiras em patas
Underwater sounds
ESCÓLIOS
The fossil group Placodermi is the most phylogenetically basal of the clade of jawed vertebrates but lacks a marginal dentition comparable to that of the dentate Chondrichthyes, Acanthodii and Osteichthyes (crown-group Gnathostomata). The teeth of crown-group gnathostomes are part of an ordered dentition replaced from, and patterned by, a dental lamina, exemplified by the elasmobranch model. A dentition recognised by these criteria has been previously judged absent in placoderms, based on structural evidence such as absence of tooth whorls and typical vertebrate dentine. However, evidence for regulated tooth addition in a precise spatiotemporal order can be observed in placoderms, but significantly, only within the group Arthrodira. In these fossils, as in other jawed vertebrates with statodont, non-replacing dentitions, new teeth are added at the ends of rows below the bite, but in line with biting edges of the dentition. The pattern is different on each gnathal bone and probably arises from single odontogenic primordia on each, but tooth rows are arranged in a distinctive placoderm pattern. New teeth are made of regular dentine comparable to that of crown-gnathostomes, formed from a pulp cavity. This differs from semidentine previously described for placoderm gnathalia, a type present in the external dermal tubercles. The Arthrodira is a derived taxon within the Placodermi, hence origin of teeth in placoderms occurs late in the phylogeny and teeth are convergently derived, relative to those of other jawed vertebrates. More basal placoderm taxa adopted other strategies for providing biting surfaces and these vary substantially, but include addition of denticles to the growing gnathal plates, at the margins of pre-existing denticle patches. These alternative strategies and apparent absence of regular dentine have led to previous interpretations that teeth were entirely absent from the placoderm dentition. A consensus view emerged that a dentition, as developed within a dental lamina, is a synapomorphy characterising the clade of crown-group gnathostomes. Recent comparisons between sets of denticle whorls in the pharyngeal region of the jawless fish Loganellia scotica (Thelodonti) and those in sharks suggest homology of these denticle sets on gill arches. Although the placoderm pharyngeal region appears to lack denticles (placoderm gill arches are poorly known), the posterior wall of the pharyngeal cavity, formed by a bony flange termed the postbranchial lamina, is covered in rows of patterned denticle arrays. These arrays differ significantly, both in morphology and arrangement, from those of the denticles located externally on the head and trunkshield plates. Denticles in these arrays are homologous to denticles associated with the gill arches in other crown-gnathostomes, with pattern similarities for order and position of pharyngeal denticles. From their location in the pharynx these are inferred to be under the influence of a cell lineage from endoderm, rather than ectoderm. Tooth sets and tooth whorls in crown-group gnathostomes are suggested to derive from the pharyngeal denticle whorls, at least in sharks, with the patterning mechanisms co-opted to the oral cavity. A comparable co-option is suggested for the Placodermi.
0 comentários:
Postar um comentário