INTRODUÇÃO À BOTÂNICA
Botânica: Scientia amabilis
Vou relatar meu encontro com a botânica. Eu sempre gostei de ciências e tive sorte de estudar sempre em escolas públicas que tinham os melhores professores tanto de ciências quanto de Biologia. Sempre que iniciava um assunto novo meus professores de Ciências da quinta à oitava série, levavam a turma para um charco que tinha ao lado da escola para observar a natureza e tomar notas no caderno do que víssemos por lá. Isso determinou em grande parte o meu gosto pelas ciencias naturais e minha decisão de fazer Biologia na Universidade Federal.
Botânica ou Fitologia, phytologia do grego: φυτεύωv, planta, é a parte da biologia que estuda a vida das plantas.
A palavra Botânica vem do grego: βοτάνη, botanē, botané, que significa planta ou erva (medicinal).
Muitas vezes a Botânica é também chamada de Biologia Vegetal ou Ciência das Plantas (Plant science).
A Botânica abrange uma ampla gama de disciplinas científicas preocupadas com o estudo das plantas, algas e fungos, incluindo a classificação, reprodução, a estrutura, o desenvolvimento, a genética, adaptação, evolução além das doenças das plantas (fitopatologia) a influência dos vegetais na cultura humana (etnobotânica). Embora os fungos e as algas sejam estudados pela botânica esses organismos não pertencem ao reino Archaeplastidae (Primolantae).

Sequência estratigráfica dos principais pólen de angiosperma
e tipos de folhas no Grupo Potomac do leste da América do Norte.
https://youtu.be/3OCcGS-zRc8
CARACTERÍTICAS GERAIS DOS VEGETAIS
CARACTERÍSTICAS MORFOFISIOLÓGICAS
Clado ARCHAEPLASTIDA

Archaeplastida ou Primoplantae


Clado Archaeplastida
1) Archaeplastida
Archaeplastida ou Primoplantae é uma das grandes linhagens de eucariotos, que compreende as plantas terrestres (embriófitas), as algas verdes (clorfófitas e carófitas) e algas vermelhas (rodófitas), e um pequeno grupo chamado Glaucophyta.
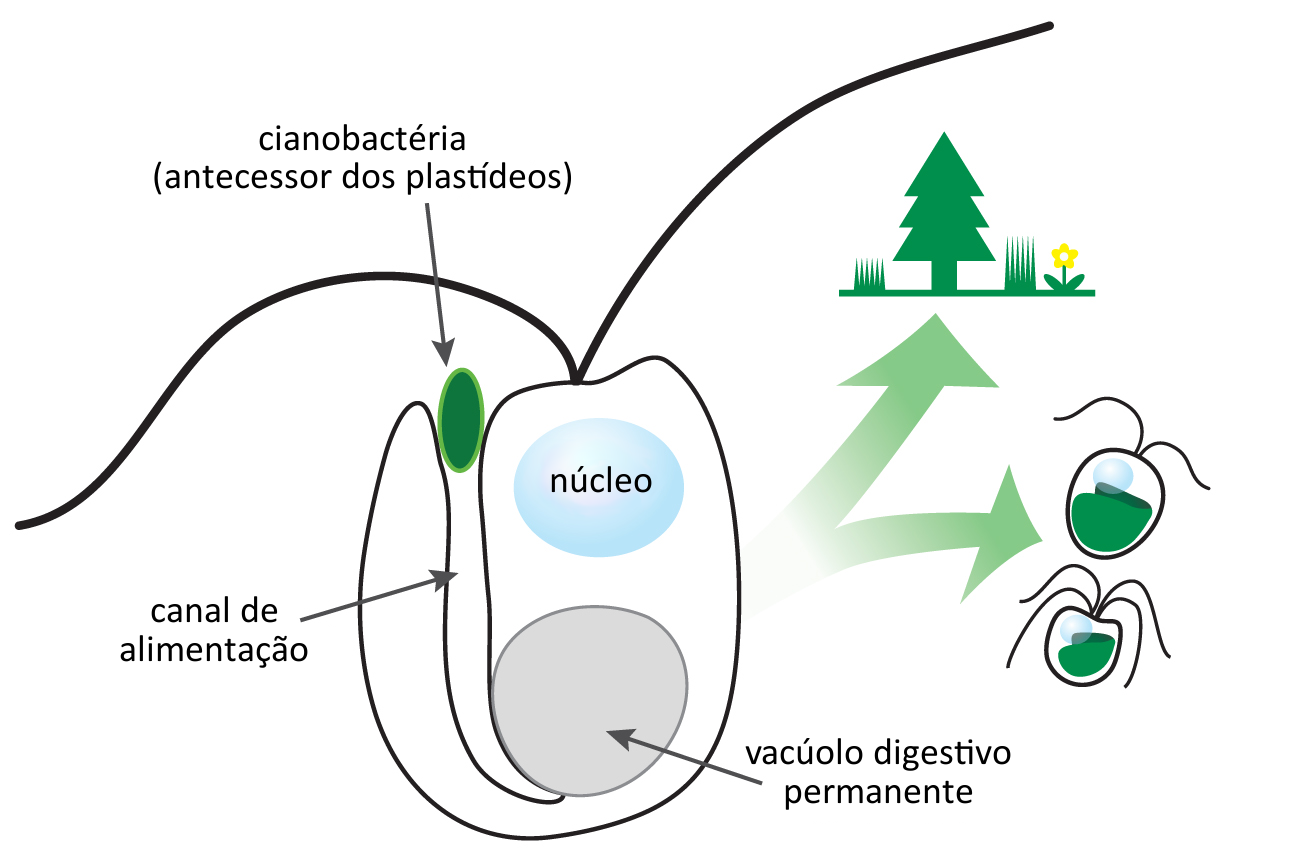
Todos estes organismos possuem plastídios rodeados por duas membranas, sugerindo que se desenvolveram diretamente a partir de cianobactérias endossimbióticas (endossimbiose primária). (Sinapomorfia: são características derivadas compartilhadas, no caso a endossimbiose primária).
O reino (supergrupo) Archaeplastida ou Primoplantae é um grupo monofilético, formado por organismos que possuem células com plastídios fotossintéticos derivados de uma endossimbiose primária, envolvendo, portanto, um procarioto fotossintetizante (cianobactéria) englobado por um eucarioto.
Nesse clado, seus representantes possuem, majoritariamente, plastídios revestidos por duas membranas, mas em algumas espécies eles foram reduzidos ou perdidos secundariamente.
Além dessa sinapomorfia, possuem o amido como reserva energética; suas mitocôndrias apresentam cristas planas; não possuem centríolos nas células; apresentam cloroplastos com clorofila a além da clorofila b em todos os seus representantes e, geralmente, apresentam uma parede celular composta por celulose.
O termo “algas” é aplicado a uma ampla variedade de organismos aquáticos fotossintetizantes
pertencentes a diversas linhagens não diretamente relacionadas umas com as outras, e, portanto,
não formam um grupo monofilético (Judd et al., 2008), todavia, as algas constantes do clado arqueplastida são fortemente unidas pelo evento fundador ancestral, e por isso não podem ser segregadas desse grupo, já que estão unidas pelo evento fundador do clado: a endossimbiose primária.

Representação esquemática da evolução do cloroplasto através da Endossimbiose Primária. Chl a: clorofila a, Chl b: clorofila b, PB: ficobiliproteínas, TLC: Transferência lateral de genes. Modificado de Bellorin & Oliveira (2006) (LOPEZ et alii, 2013).
Endossimbiosse ontem e hoje
A principal evidência de que os Archaeplastida formam um grupo monofilético vem de estudos genéticos e moleculares (cpDNA), que indicam que seus plastídeos provavelmente tem uma origem única; seriam o resultado de um único evento de endossimbiose primária de uma cianobactéria. Essa cianobactéria teria invadido (ou foi engolfada) uma célula de um eucarioto unicelular heterotrófico, e com o passar do tempo tornou-se o cloroplasto.

Hipótese endossimbiótica na origem dos grandes grupos taxonômicos.

Análise do cpDNA corrobora a hipótese monofilética para os arqueplastidas.
As células dos Archaeplastida normalmente não têm centríolos e têm mitocôndrias com cristas planas. Eles geralmente têm uma parede celular que contém celulose e armazenam amido como carboidrato de reserva.
Todavia, essas características são compartilhadas com outros eucariotos.
Os arqueplastidas dividem-se em duas linhas evolutivas principais. A primeira inclui as algas vermelhas, que apresentam clorofila a como pigmento fotossintetizante e ficobiliproteínas (ficobilinas), como a maioria das cianobactérias, e armazenam amido no citoplasma (amido das florídeas).
Uma segunda linha evolutiva inclui as algas verdes e as plantas terrestres, conhecidas como Viridiplantae (do latim: plantas verdes) ou Chloroplastida, apresentam clorofilas a e b, e carotenóides, mas não possuem ficobiliproteínas, e o amido é acumulado dentro dos cloroplastos.


Clorofila a e b (STREIT et alii, 2005)




Clorofila a e b comparada com bacterioclorofila

Absorção das clorofilas

Clado Viridiplantae




Clado Archaeplastida

Relações filogenéticas das briófitas sensu lato.
Modificado de Bowman et al. (2017).
2) Plantas terrestres ou Embryophytas
Plantas terrestres ou embriófitas (grupo de organismos cuja característica em comum é a presença de um embrião multicelular matrotrófico, um sinônimo para plantas terrestres), são organismos eucariotos, multicelulares, clorofilados, com clorofila a e como pigmento acessório a clorofila b e β-caroteno (carotenóides), que surgiram como resultado de uma endossimbiose primária de uma cianobactéria por um organismo unicelular eucarioto heterotrófico. Apareceram no registro fóssil por volta de 425 milhões de anos (período Siluriano superior ou até mesmo no Ordoviciano, portanto no paleozóico).

Os carotenóides (Figura acima) são geralmente tetraterpenóides de 40 átomos de carbono, de coloração amarela, laranja ou vermelha. São encontrados em vegetais e classificam-se em carotenos ou xantofilas. Os carotenos são hidrocarbonetos poliênicos com variados graus de insaturação, e as xantofilas são sintetizadas a partir dos carotenos, por meio de reações de hidroxilação e epoxidação. O b-caroteno e o licopeno são exemplos de carotenos, enquanto a luteína e a zeaxantina são xantofilas (Ambrósio e cols, 2006).

3) Endossimbiose primária, clorofila a e b e carotenoides, amido no cloroplasto, cloroplasto com tilacoides, gametas biflagelados, celulose na parede celular.

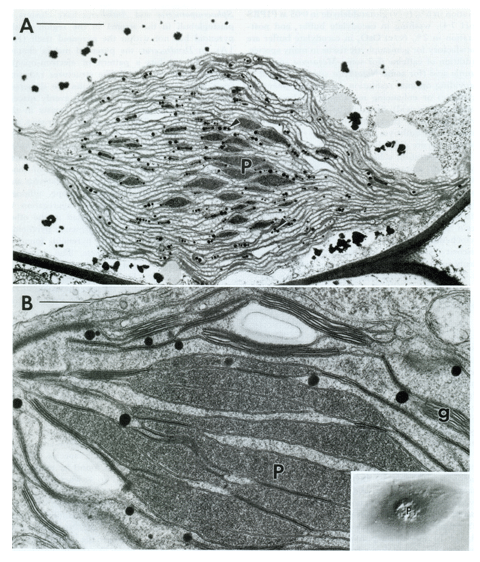
(A) Micrografia eletrônica de um cloroplasto de Sphaerosporoceros adscendens, um cloroplasto de antocerófita bastante "típico". As subunidades do pirenoide múltiplo (P) são mais eletro-opacas do que o estroma circundante. Os tilacóides de canal conectam pilhas de grana de ponta a ponta. Numerosos plastoglóbulos (setas) são esferas elétron-opacas encontradas no estroma. S, amido (starch); barra, soma 50. (B) Micrografia eletrônica da região pirenóide de Phaeoceros laevis. As subunidades dos pirenóides (P) são mais eletro-opacas do que o estroma circundante. O pirenóide é dissecado pelo estroma, bem como pelos tilacóides organizados em ambos grana (g) e lamela simples (estroma). Barra, 05 sum. Detalhe: micrografia de interferência diferencial de Nomarski de um cloroplasto de Anthoceros punctatus com um pirenoide múltiplo proeminente (P), x 200. (VAUGHN, 1992).
[(A) Electron micrograph of a chloroplast of Sphaerosporoceros adscendens, a rather 'typical' anthocerote chloroplast. Subunits of the multiple pyrenoid (P) are more electron-opaque than the surrounding stroma. Channel thylakoids connect grana stacks together end-to-end. Numerous plastoglobuli (arrows) are electron-opaque spheres found in the stroma. S, starch; bar, 50 sum. (B) Electron micrograph of the pyrenoid region of Phaeoceros laevis. The pyrenoid subunits (P) are more electron-opaque than the surrounding stroma. The pyrenoid is dissected by stroma as well as thylakoids organized into both grana (g) and single (stroma) lamellae. Bar, 05 sum. Inset: Nomarski differential interference micrograph of an Anthoceros punctatus chloroplast with a prominent multiple pyrenoid (P), x 200. (VAUGHN, 1992)].

Antócero (geocities)
4) Apresentam metagênese ou alternância de gerações
Tipo de reprodução onde se alternam a reprodução sexuada (esporofítica, diploide) e assexuada (gametofítica, haploide) como fases distintas de seu desenvolvimento ou ciclo de vida.


Onde são produzidos os gametas (anterozóide e oosfera)
Fase esporofítica
Onde os gametas ao se fundirem originam o zigoto ou esporófito. Ao se desenvolver o esporófito (através de mitoses sucessivas) produz o esporófito adulto quando passa a produzir esporos, através de meiose espórica.
5) Habitat
São organismos adaptados a vida em terra firme, embora algumas possam viver na água.



6) São Autótrofos


Ciclo de vida dos musgo (simplificado) grupo basal das embriófitas.

Ciclo de vida de uma monilófita um clado monofilético (simplificado).
As plantas são capazes, através da fotossíntese, de usarem a luz do sol, água, sais minerais e o gás carbônico da atmosfera e convertê-los em glicose (um carboidrato simples) e a partir deste todos os outros compostos e no final produzem oxigênio. As produzem seu próprio alimento, sendo portanto autotróficas.
7) Embrião multicelular
São também chamadas de Embriófitas, por possuírem embriões multicelulares (maciços sem cavidades), sinapomorfia das plantas terrestres, envolvidos por tecido de origem materna (retidos no gametófito feminino, onde são protegidos e alimentados). O esporófito cresce e se desenvolve com um parasita do gametófito nas marcantiófitas, musgos e antocerófitas.
8) Matrotrofia
8) Matrotrofia
Apresentam matrotrofia, i.e., o embrião dos metáfita é retido e nutrido pelo organismo materno (gametófito feminino), o que os diferenciam das algas, e esta característica é uma vantagem adaptativa para a vida em terra (fora da água, lembre-se que as plantas terrestres evoluíram das algas Carófitas (algas verdes). Esta característica juntamente com a anterior é a sinapomorfia do clado Embryophyta. Todas os organismos que apresentam as características acima pertencerem ao clado Archaeplastida (Plantae).
PLANTAS TERRESTRES
EMBRYOPHYTA
As plantas terrestres podem ser dividido em dois grupos informais:
Grupo das plantas avasculares (sem vasos condutores de seiva): Bryophyta
Grupo das plantas vasculares (com vasos condutores de seiva): Traqueophyta.
Grupo das plantas vasculares (com vasos condutores de seiva): Traqueophyta.
As plantas traqueófitas (vasculares) são divididas informalmente em:
plantas sem sementes (Criptógamas):
plantas sem sementes (Criptógamas):
"Pteridophytas"
plantas com sementes (Espermatófitas):
Cicadophyta, Ginkgophyta, Gnetophyta, Conipherophyta e Angiospermae
As plantas Espermatófitas podem ser divididas por sua vez em:
Plantas com semente nua: Cicadofita, Ginkgofita, Coniferofita, Gnetofita ("Gimnospermas") e
Plantas com flores e frutos: Anthophyta ("Angiospermas"): Monocotiledôneas e Eudicotiledôneas.
Plantas com semente nua: Cicadofita, Ginkgofita, Coniferofita, Gnetofita ("Gimnospermas") e
Plantas com flores e frutos: Anthophyta ("Angiospermas"): Monocotiledôneas e Eudicotiledôneas.

A diversidade de cloroplastos de Anthocerophyta. (A) O modelo de antócero Anthoceros agrestis. (B) Nothoceros vincentianus tem um único cloroplasto por célula e um pirenóide proeminente (p). (C) Nothoceros fuegiensis tem vários cloroplastos por célula e não tem pirenóide. (D) O pirenóide no cloroplasto de Notothylas breutelii, mostrado como as unidades elétron-densas atravessadas pelos tilacóides. (E) O cloroplasto sem pirenóide de Leiosporoceros dussii, com grana (g) e grão (s) de amido, mas sem pirenóide. Reproduzido, com permissão, de J.C. Villarreal (B, C, E) e K. Renzaglia (D). (WEI-LI and VILLAREAL, 2017)


Cladograma mostrando os taxa que compõem o clado Embryophyta
(plantas terrestres), mostrando as características sinapomórficas desse grupo.
Embrião multicelular maciço retido no gametófito feminino
Capacidade de produzir cutícula (cutina): a cutina é uma macromolécula, um polímero formado por muitos ácidos graxos de cadeias longas, ligados uns aos outros por ligações ésteres, constituindo uma rede tridimensional rígida.
É uma macromolécula formada por ácidos graxos de cadeia longa com grupos alcoólicos ligados entre si por ligações éster, formando assim um polímero amorfo de tamanho e peso molecular indefinidos. Mais precisamente, é um poliéster natural. As duas famílias de ácidos graxos que ele contém são tipicamente lipídios de cadeia C16 saturados (ácido palmítico) e lipídios de cadeia C18 saturados (ácido esteárico) ou monoinsaturados (ácido oleico). As paredes externas das células epidérmicas são incrustadas com cutina.
Como vimos acima a cutina é uma substância de constituição lipídica que pode estar impregnada na parede celular ou então formando uma camada na superfície externa dessas células. A principal função da cutícula é proteger contra a perda excessiva de água e contra os raios UV.

A

B
(A) Esquema da estrutura da cutícula, a cobertura protetora da epiderme das folhas e de caules jovens no estádio de folhas completamente expandidas. (B) Eletromicrografia da cutícula de uma célula glandular de uma folha jovem (Lamium sp.), mostrando a presença de camadas cuticulares indicadas em A, com exceção das ceras epicuticulares, as quais não são visualizadas. (51.000x) (A, segundo Jeffree, 1996; B, de Gunning e Steer, 1996).

Em (A) vemos a cutina

Cutina no tomate
Amido como principal reserva
Esporos com paredes de esporopolenina
Anterídios e arquegônios com camadas de células estéreis
Clorofila a, clorofila b e carotenóides
Plasmodesmos
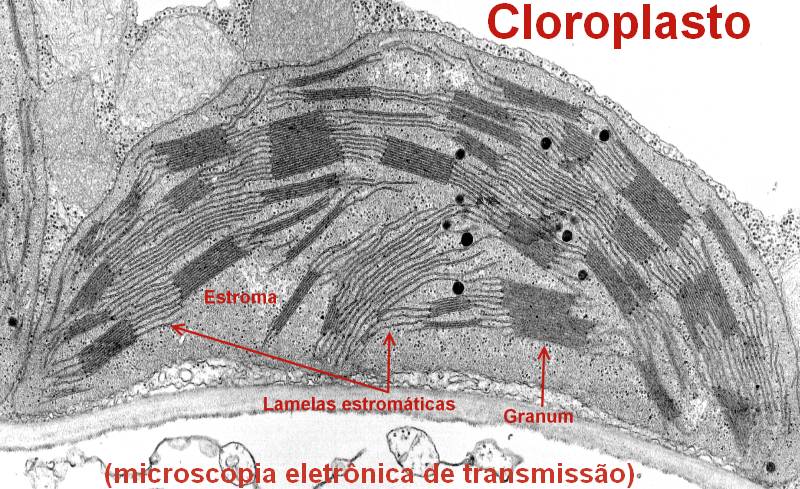
Tilacoides com grana
Metagênese
Saída do ambiente aquático, Xiloglicano e Evolução das plantas
O xiloglucano (XyG) é o principal polissacarídeo da matriz não celulósica, não péctica da parede celular da maioria das plantas terrestres. Inicialmente acreditava-se que estava restrito às plantas terrestres, genes de síntese e modificação/degradação para XyG foram detectados em algas verdes carofíceas (Chariophycean green algae CGA).
Recentemente, uma função totalmente nova do XyG foi descoberta como um potente agregador de solo liberado por raízes e rizóides de todos os principais grupos de plantas terrestres (Del Bem, 2018).
Embryophyta é um grupo monofilético que é filogeneticamente mais relacionado a dois grupos de CGA: Zygnematophyceae e Coleochaetophyceae (Laurin-Lemay et al., 2012). (Del Bem, 2018).
Muitos traços comuns foram encontrados em plantas terrestres e carófitas, um dos quais é o polissacarídeo hemicelulósico da parede celular XyG (Ikegaya et al., 2008; Sorensen et al., 2010, 2011; Herburger et al., 2018).
XyG é o principal polissacarídeo de matriz não celulósica não péctica nas paredes celulares da maioria das plantas terrestres e
consiste em um esqueleto de D-glucana ligada por ligações β (1 → 4) que é posteriormente substituída por resíduos de xilosil. Os resíduos de xilosil podem ser ainda substituídos por outros glicosil (principalmente galactosil e fucosil) e resíduos não glicosil que podem variar entre os grupos de plantas (Pauly & Keegstra, 2016). Uma unidade típica de XyG é mostrada na Fig. abaixo (a) com as enzimas associadas necessárias para sua síntese, modificação e degradação.(Del Bem, 2018).

Xyloglucan (XyG)-related genes and their presence across charophycean green algae (CGA) lineages.
(a) A typical XyG subunit (XXFG) showing its molecular structure and the enzymes required for XyG synthesis (glycosyl transferases, green), degradation (glycosyl hydrolases, red) and endohydrolysis/transglycosylation (XTH, blue).
(b) The number of XyG-related genes detected by Blastp (e-value < e−20) in Klebsormidium nitens NIES-2285 genome v.1.0
(http://www.plantmorphogenesis.bio.titech.ac.jp/~algae_genome_project/klebsormidium/). Homologs of AXY8 were not detected (ND) even with an e-value cutoff of e−4.
(c) The number of CGA genera with positive Blastp hits (e-value < e−20) in transcriptomic data for each XyG-related gene. The assembled transcriptomes were obtained from the OneKP project (http://www.onekp.com/public_data.html) for the genera Penium, Cosmarium, Staurodesmus, Phymatodocis, Bambusina, Pleurotaenium, Micrasterias, Gonatozygon, Staurastrum, Euastrum, Onychonema, Xanthidium, Desmidium, Roya, Mesotaenium, Spirotaenia, Zygnema, Zygnemopsis, Nucleotaenium, Cylindrocystis, Netrium, Closterium, Mougeotia, Planotaenium and Spyrogira (Class Zygnematophyceae); Coleochaete and Chaetosphaeridium (Coleochaetophyceae); Chara (Charophyceae); Klebsormidium and Entransia (Klebsormidiophyceae); and Mesostigma (Mesostigmatophyceae). From the NCBI EST database (https://www.ncbi.nlm.nih.gov/nucest/?term=chlorokybus) for the genus Chlorokybus (Chlorokybophyceae; 23 716 unique sequences) and the NCBI Sequence Set Browser (https://www.ncbi.nlm.nih.gov/Traces/wgs/) for the genera Coleochaete (Coleochaetophyceae, accession GBSL01) and Nitella (Charophyceae, accession GBST01). *, Classes composed of a single genus.
XyG was first thought to be absent from CGA (Popper & Fry, 2003) and primarily involved in the mechanical properties of the cell walls of eudicots and noncommelinid monocots by interacting with cellulose (Thompson & Fry, 2000). This view of XyG was broadened and somewhat challenged during the last decade. Ikegaya et al. (2008) detected a XyG-like polysaccharide in the CGA Spyrogira (Zygnematophyceae) and proposed a role for it in cell–cell attachment. Cavalier et al. (2008) were able to produce Arabidopsis plants deficient in XyG by disrupting two XyG xylosyltransferases (XXT1 and XXT2). The double mutant xxt1 xxt2 displayed only minor phenotypic alterations, suggesting that XyG is not essential for the fundamental mechanical properties of cell walls. We wrote a paper during my PhD (Del Bem & Vincentz, 2010) suggesting a model for the emergence of XyG-related genes showing that two essential enzymes in XyG synthesis and modification – β-Glucan synthase from Cellulose Synthase-Like subfamily C (CSLC) and XyG endotransglucosylase/hydrolase (XTH) – most likely originated in charophytes, before the emergence of embryophytes. Now new work (Galloway et al., 2018) has extended the understanding of XyG, showing that it is a potent soil particle aggregator released by the roots and rhizoids of all major groups of embryophytes. They detected XyG in a variety of soils suggesting that it is involved in soil formation and its properties.
Here I extend this idea a little further by proposing that XyG originated during the terrestrialization of early Streptophyta as a key adaptation, allowing unicellular CGA to interact and modify early soils and substrates.

Modelo para a emergência da maquinaría genética do xiloglucano (XyG) em Streptophyta. As caixas separam os principais grupos de Viridiplantae com base na presença de conjuntos particulares de genes. Todas as enzimas responsáveis pela síntese de XyG surgiram no último ancestral comum (LCA) de Klebsormidiophyceae, Charophyceae, Coleochaetophyceae, Zygnempatophyceae e Embryophyta. Os homólogos da glicosil hidrolase XYL1/AXY3 e β-Gal10 originaram-se antes do surgimento da maquinaria sintética de XyG e provavelmente foram posteriormente cooptados para a degradação de XyG. Os homólogos da α-fucosidase AXY8 surgiram posteriormente na LCA de Coleochaetophyceae, Zygnematophyceae e Embryophyta. O esquema de cores para a função enzimática na síntese, degradação e modificação de XyG são os mesmos da Fig. 1. As relações filogenéticas entre os grupos Viridiplantae são baseadas em Turmel et al. (2007), Timme et al. (2012) e Wickett et al. (2014). *, Grupos com espécies terrestres ou terrestres facultativas.
[Model for the emergence of xyloglucan (XyG) genetic machinery in Streptophyta. The boxes separate the major Viridiplantae groups based on the presence of particular sets of genes. All enzymes responsible for XyG synthesis emerged in the last common ancestor (LCA) of Klebsormidiophyceae, Charophyceae, Coleochaetophyceae, Zygnempatophyceae and Embryophyta. Homologs of the glycosyl hydrolase XYL1/AXY3 and β-Gal10 originated before the emergence of XyG synthetic machinery and were most likely later co-opted for XyG degradation. The homologs of the α-fucosidase AXY8 emerged later in the LCA of Coleochaetophyceae, Zygnematophyceae and Embryophyta. The color scheme for enzyme function in XyG synthesis, degradation and modification are the same as those in Fig. 1. The phylogenetic relationships between Viridiplantae groups are based on Turmel et al. (2007), Timme et al. (2012) and Wickett et al. (2014). *, Groups with terrestrial or facultative terrestrial species.] Continua aqui.
Hepaticas, Antoceros e Musgos
Marchantiophytas,
Anthocerophytas e
Bryophyta
De maneira geral, as briófitas caracterizam-se por seu tamanho pequeno, podendo variar de poucos milímetros até um metro, como ocorre com alguns musgos de hábito pendente e presença de gametófitos talosos ou folhosos, clorofilados e fotossintetizantes (Lemos-Michel 2001). As Bryophytas apresentam parede celular celulósica e, algumas vezes, cutícula com cutina e alternância de gerações bem definida.
A geração sexuada e diplóide é representada por um esporófito de vida efêmera, não ramificado, dependente e aderido ao gametófito (geração assexuada), que é haplóide, fotossintetizante, perene e livre, produzindo os órgãos sexuais: anterídios e arquegônios (Schofield 1985).
São abundantes em ambientes úmidos e sombrios, no interior de matas ou são típicas de áreas urbanas (Lemos-Michel 2001). Também ocorrem em hábitats aquáticos (Delgadillo & Cárdenas 1990) e em lugares secos e desérticos, já que são adaptadas à dessecação e sobrevivem como esporos dormentes ou gametófitos secos (Margulis & Schwartz 2001). Por tolerarem condições ambientais extremas estão amplamente distribuídas pelo mundo, desde ambientes árticos, tundra e florestas boreais até florestas tropicais, ambientes submersos, cavernas escuras e rochas expostas, tolerando também aspersão de água salgada, mas nunca sendo encontradas em ambiente marinho (Delgadillo & Cárdenas 1990, Frahm 2003). As briófitas desenvolvem-se nos mais diversos substratos: troncos vivos ou em decomposição, ramos, húmus, superfície de rocha, solos calcários, argilosos ou arenosos, folhas vivas e outros materiais orgânicos. Também podem ocorrer sobre telhados, muros e calçadas, além de substratos não usuais, como cascos de tartaruga e calçados velhos (Frahm 2003) (modif. de Bordim, (2008)).
A transição das primeiras plantas, do ambiente aquático, para o ambiente terrestre ocorreu aproximadamente há 470 milhões de anos atrás.
O aparecimento de importantes sinapomorfias foi essencial para a conquista desse ambiente pelas embriófitas, garantindo a relativa independência do habitat aquático.
Entre essas novidades evolutivas, o surgimento de uma camada extracelular lipofílica contínua recobrindo as partes aéreas das plantas, a cutícula vegetal, foi crucial para o sucesso evolutivo desse grupo.

Organização da cutícula (Matos, 2020)

Esquema das principais funções da cutícula vegetal nas plantas. (A): Reduzir a perda de água e difusão de gases. (B): Evitar a acumulação de água e propiciar autolimpeza da superfície foliar. (C): Mediar a interação planta-inseto. (D): Prevenir a penetração de patógenos via sinalização. (E): Auxiliar na termorregulação e proteger contra os raios UV. (F): Promover suporte mecânico. Retirado de Báez-Sañudo et al. (2013).(Matos, 2020).

Diferentes morfologias das ceras epicuticulares obtidas por microscopia eletrônica de varredura (MEV). Morfologias amorfas (A-C). A: morfologia do tipo filme em Hydrocotyle bonariensis Lam. (Apiaceae). B: morfologia do tipo camadas fissuradas em Crassula ovata (Mill.) Druce (Crassulaceae). C: morfologia do tipo crosta em Cynanchum sarcostemma Lillo (Asclepiadaceae). Morfologias cristaloides (D-I). D: morfologia do tipo plaquetas no musgo Polytrichadelphus magellanicus (Hedw.) Mitt. (Polytrichaceae). E: morfologia do tipo túbulos de nonacosan-10-ol em Nelumbo nucifera Gaertn. (Nelumbonaceae). F: morfologia do tipo lâminas ordenadas em paralelo em Convallaria majalis L. (Convallariaceae). G: morfologia do tipo grânulos em Aegiceras corniculatum (L.) Blanco (Aegicerataceae). H: morfologia do tipo bastonetes enrolados em Buxus sempervirens L. (Buxaceae). I: sintopismo de plaquetas e lâminas em Benthamia alyxifolia (F. Muell. Ex Benth.) Tiegh. (Loranthaceae). Imagens retiradas e modificadas de Clayton-Greene et al. (1985), Barthlott et al. (1998; 2017) e Koch et al. (2009a).(Matos, 2020).
A cutícula é composta pela cutina a qual é envolvida pelas ceras cuticulares, e desempenha um papel multifuncional nas plantas, sobretudo reduzindo a perda de água não estomática e, portanto, evitando a dessecação, além de atuar protegendo as células contra os raios nocivos do sol (UV). Além disso produz corpos oleosos no interior de células com diversas funções ainda não esclarecidas (deterrência de herbívoros, proteção contra fungos e bactérias, proteção contra dessecação, manutenção do equilíbrio osmótico, e proteção contra UV). (Deterrência: ação de desencorajar, dissuadir, retardar ou impedir a ação de herbívoros).

A maior parte da diversidade química de hepática reside dentro das células do corpo oleoso, uma sinapomorfia da linhagem (presença de corpos oleosos no interior das células). Romani, Banic et al. (2015) mostram que os órgãos oleosos defendem a planta contra artrópodes. Além disso, a diferenciação das células do corpo oleoso requer um fator de transcrição MpC1HDZ, que mostra altos níveis de expressão gênica antes da diferenciação do corpo oleoso (Romani e cols. 2015).
As ceras são formadas por diversos compostos hidrofóbicos acíclicos de cadeia longa tais como, os aldeídos, álcoois primários e secundários, alcanos, cetonas e ésteres.
Também são encontrados compostos cíclicos como triterpenos e esteroides. Na superfície externa da camada cuticular, as ceras epicuticulares podem formar diferentes tipos de ornamentos, variando desde amorfas a cristaloides.
Essas plantas são consideradas as pioneiras na transição do ambiente aquático para o terrestre. E são frequentemente comparadas aos anfíbios que também fizeram essa transição da água para a terra firme, mas ainda se mantém dependentes da água.
São avasculares, "criptógamas" e possuem um ciclo de vida com a geração gametofítica dominante. Essas plantas podem tolerar condições ambientais extremas e por isso estão amplamente distribuídas no mundo, ocorrendo desde o Ártico até as florestas tropicais, sendo mais abundantes em ambientes úmidos. Elas compõem o segundo maior grupo de plantas terrestres.
No mundo, são conhecidas aproximadamente 20.000 espécies e, no Brasil, cerca de 1.524, ocorrendo em todas as regiões e biomas brasileiros, inclusive na Caatinga, sendo a Mata Atlântica do Sudeste o centro de maior diversidade dessas plantas.
O termo “briófitas” é utilizado para designar três divisões distintas: Marchantiophyta (hepáticas), Bryophyta (musgos) e Anthocerotophyta (antóceros).
Outrora agrupadas sob o nome de briófitas, hoje reconhecidas com três divisões distintas, cada uma com seu ancestral comum.
Todas as três são plantas avasculares, i.e., não possuem vasos condutores de seiva (xilema e floema).
São consideradas como intermediárias entre as algas verdes e as plantas vasculares.
Elas compartilham algumas características com as algas, como, por exemplo, o cloroplasto.
Porém, diferentemente destas, as briófitas já apresentam adaptações para a vida fora da água, constituindo-se em plantas da transição do ambiente aquático para o terrestre.
Para viver no meio terrestre são necessárias adaptações às condições desse novo meio. Uma delas é evitar a perda de água pela evaporação que ocorre na superfície da planta em contato com o ar. Para isso, as plantas terrestres apresentam, entre outras adaptações, um revestimento protetor sobre a epiderme, chamado cutícula, com cutina, uma cera impermeabilizante protetora, produzidas unicamente pelas células da epiderme.
A cutina é uma macromolécula, componente principal da cutícula das plantas terrestres. Trata-se de um polímero formado por muitos ácidos graxos de cadeia longa, que estão unidos uns aos outros por ligações éster, criando uma rede rígida tridimensional, na superfície da epiderme. A cutina é formada por ácidos graxos 16:0 e 18:1 (isto é, de 16 átomos de carbono sem ligações duplas, e de 18 átomos de carbono com uma dupla ligação cis). A cadeia pode ter grupos hidroxila ou epóxido, interiormente ou no extremo oposto ao grupo carboxila.

Cutícula com cutina secretada pela epiderme
Funciona como uma proteção contra perda dágua (desidratação), proteção contra infecção (entrada de patógenos) e contra traumas mecânicos, alem de atuar minimizandoos efeitos nocivos da radiação solar UV.
A primeira etapa da via de biossíntese para a formação dos ácidos graxos de cadeias muito longas (VLCFAs) cuticulares, ocorre com a biossíntese de novo das cadeias acil C16 (palmitato) pelos cloroplastos no mesofilo, e é concluída com a extensão dessas cadeias no retículo endoplasmático das células da epiderme.
A função primária da cutícula da planta é como uma barreira de permeabilidade à água (i.e., evitando a evaporação da água da superfície epidérmica) e também impedindo que água externa e solutos entrem nos tecidos.
Em adição a sua função como barreira à permeabilidade para água e outras moléculas (evitando a perda d´água) a micro e nano estrutura da cutícula possuiem propriedades especializadas de superfície que evitam a contaminação dos tecidos vegetais com água externa, sujeira e microorganismos.
Os órgãos aéreos de muitas plantas, como as folhas do lótus sagrado (Nelumbo nucifera), possui propriedades ultra-hidrofóbicas e autolimpantes que foram descritas por Barthlott e Neinhuis (1997). O efeito-lótus tem aplicações em materiais técnicos biomiméticos.

Micrografias eletrônicas de varredura da superfície foliar adaxial de superfícies lisas, molháveis (a-d) e ásperas, repelentes à água (e-h). As folhas lisas de Gnetum gnemon (a) e Heliconia densiflora (b) são quase completamente desprovidas de microestruturas, enquanto as de Fagus sylvatica (c) e Magnolia denudata (d) são caracterizadas por nervatura rebaixada e elevada, respectivamente. As superfícies rugosas de Nelumbo nucifera (e) e Colocasia esculenta (f) são caracterizadas por células epidérmicas papilosas e uma camada adicional de ceras epicuticulares. As folhas de Brassica oleraces (g) são densamente cobertas por cristalóides de cera sem serem papilosas, e as superfícies das pétalas de Mutisia decurrens (h) são caracterizadas por dobras cuticulares. Barras: 100 µm (a-d) e 20 µm (e-h). (Barthlott e Neinhuis, 1997).
[Scanning electron micrographs of the adaxial leaf surface of smooth, wettable (a-d) and rough, water repellent (e-h) leaf surfaces. The smooth leaves of Gnetum gnemon (a) and Heliconia densiflora (b) are almost completely lacking microstructures while those of Fagus sylvatica (c) and Magnolia denudata (d) are characterized by sunken and raised nervature, respectively. The rough surfaces of Nelumbo nucifera (e) and Colocasia esculenta (f) are characterized by papillose epidermal cells and an additional layer of epicuticular waxes. Brassica oleraces leaves (g) are densely covered by wax crystalloids without being papillose, and the petal surfaces of Mutisia decurrens (h) are characterized by cuticular folds. Bars: 100 µm (a-d) and 20 µm (e-h). (Barthlott and Neinhuis, 1997).]

O desenho resume a conexão entre rugosidade e autolimpeza. Enquanto em superfícies lisas as partículas são redistribuídas principalmente pela água (a), elas aderem às superfícies das gotículas em superfícies ásperas e são removidas das folhas quando as gotículas rolam (b).
[The drawing summarizes the connection between roughening and self-cleaning. While in smooth surfaces the particles are mainly redistributed by water (a), they adhere to the droplets surfaces in rough surfaces and are removed from the leaves when the droplets roll off (b).]

Fig. 5. Partícula contaminante em uma superfície de asa regularmente esculpida de Cicada orni, demonstrando a diminuição da área de contato entre uma partícula e uma superfície rugosa. Barra: 1µm. Fig. 6. Gota de mercúrio na superfície epidérmica adaxial papilosa de Colocasia esculenta demonstrando o efeito da rugosidade na molhabilidade. Devido à diminuição da área de contato entre o líquido e a superfície, o ar fica preso entre a gota e a folha, resultando em uma superfície repelente à água particularmente forte. Barra: 20 µm. Fig. 7. Gota de mercúrio na superfície foliar adaxial de Colocasia esculenta demonstrando o Efeito Lótus. As partículas contaminantes aderem à superfície da gota e são removidas da folha enquanto a gota rola. Barra: 50 µm.
[Fig. 5. Contaminating particle on a regularly sculptured wing surface of Cicada orni, demonstrating the decreased contact area between a particle and a rough surface. Bar: 1µm. Fig. 6. Mercury droplet on the papillose adaxial epidermal surface of Colocasia esculenta demonstrating the effect of roughness on wettability. Due to the decreased contact area between liquid and surface, air is enclosed between the droplet and the leaf resulting in a particularly strong water repellent surface. Bar: 20 µm. Fig. 7. Mercury droplet on the adaxial leaf surface of Colocasia esculenta demonstrating the Lotus-Effect. Contaminating particles adhere to the surface of the droplet and are removed from the leaf while the droplet rolls off. Bar: 50 µm.]
The biological implications of the Lotus-Effect. The cuticle is the outermost barrier of plants towards their environment and is, therefore, the first protective layer (Dickinson 1960; Martin 1964; Campbell et al. 1980; Juniper 1991). Because the air contains many kinds of particles, leaf surfaces are continuously contaminated. Many deposits are ± neutral, but various kinds of contamination may cause considerable damage to the plants, depending on size and chemical nature. It was shown that in polluted areas where plants are heavily contaminated with dust, leaf surface temperatures increased under insolation (Eller 1977). In addition, particles within a certain size range may occlude stomata and influence stomatal diffusive resistance (Fl�ckiger et al. 1979). These and other interactions between dust particles and plants have been reviewed extensively by Farmer (1993). Water repellent plants escape from those harmful effects through Lotus-Effect. Although it was shown that particles are captured more effectively by rough leaf surfaces (Chamberlain 1967; Belot and Gauthier 1975), this disadvantage is compensated for by a very effective self-cleaning capability.
The Lotus-Effect plays another important role in the defense against pathogens. Spores and conidia of pathogenic microorganisms, as well as inorganic particles, are deposited on the leaf surfaces. Again, wettability is important for the adhesion of microorganisms to leaf surfaces (Rogers 1979). In addition, on water repellent surfaces, spores and conidia are deprived of the water necessary for germination (Campbell, et al. 1980; Allen et al. 1991; Juniper 1991). Therefore, the epicuticular wax crystalloids and their physical properties may be regarded as the first line of defense against pathogens. Our results indicate that the Lotus-Effect may be the most important function of epicuticular waxes and the reason for pervasive microsculpturing of many leaf surfaces. There are but a few pathogens that are able to overcome this barrier. Powdery mildews, for example, contain a small amount of water within their conidia which enables them to germinate on virtually dry surfaces. A dry surface seems to be beneficial to them, while there is some indication that a wet surface may impede germination (Wheeler 1981).
As shown before, epicuticular wax crystalloids provide a highly effective self-cleaning surface to many plants. On the other hand, they are very fragile structures and may be easily altered, especially by mechanic abrasion (van Gardingen et al. 1991; Bermadinger-Stabentheiner 1994). This also influences the water repellent function: within altered areas, particles may be retained permanently. If the damage is not too grievous, it can be compensated for by the regeneration of the wax crystalloids (e.g. Hallam 1970). Apart from naturally occurring wax alterations, some anthropogenic influences can be very serious. This is especially true for surfactants. They are an important component in all-water based pesticides, since they enable the uptake of an active ingredient through the cuticle (Stevens and Bukovac 1985; Lownds et al. 1987; Knoche and Bukovac 1993). However, the surfactants cause considerable damage to wax ultrastructure (Noga et al. 1987; Wolter et al. 1988). Due to the alterations in the wax ultrastructure, the wettability of the leaves is increased for at least several days and water is retained within the altered areas. As a result, contaminating particles including spores and conidia of pathogens are also found within the areas of altered waxes (Neinhuis 1992). Under unfavourable conditions, the probability of an infection in a plant may be enhanced, which is in contrast to the aim of a pesticide application.
The Lotus-Effect is not restricted to plants; indeed, it is of an overall biological importance, e.g. for insects. Especially those insects with large wings, which cannot be cleaned by legs, have water repellent wing surfaces and exhibit the self-cleaning ability (Wagner et al. 1996). In this case, not only the removal of particles is of interest, but also the maintenance of flight capability, which may be lost due to an unequal load on the wings.
Segundo Budke, Goffinet and Jones, (2013) a proteção contra desidratação fornecida pela cutícula materna (do gametófito feminino) melhora a aptidão da prole no musgo Funaria hygrometrica e nos esporófitos de todas as plantas vasculares.

Sporophytes of Funaria hygrometrica.
(A) Estágio de manipulação, esporófito em forma de lança, esporângio não expandido, 15 − 20 mm de altura, coberto por um caliptra gametofítica (c) com região de rostro estreito (r) no ápice (Tabela 1, estágio de desenvolvimento 4); a seta indica o ápice do esporófito abaixo da caliptra. (B, C) Esporófito com esporângio totalmente expandido e anel vermelho (a) (Tabela 1, estágio de desenvolvimento 9); (B) as linhas ilustram o ângulo de inclinação do esporângio; (C) dentes peristômios vermelhos (p) visíveis através da tampa do esporângio. (D) Esporófito seco ao ar com uma região mais escura cheia de esporos visível através da parede do esporângio; as linhas ilustram a medida do diâmetro do anel. (E) Anel malformado e de menor diâmetro; o anel é não circular, penetrando no centro da tampa do esporângio. (F) Malformação do peristômio com alguns dentes do peristômio fundidos ao interior da tampa do esporângio; deiscência de esporângio incapacitante. (G) Tampa de esporângio naturalmente deiscente com tampa agora ausente; revelando dentes peristômios maduros e devidamente formados. Barras de escala: (A, B, D) = 1 mm; (C, E, G) = 0,5 mm; (F) = 0,25 mm. (Budke, Goffinet e Jones, 2013)
[(A) Manipulation stage, spear-shaped sporophyte, sporangium unexpanded, 15 − 20 mm tall, covered by a gametophytic calyptra (c) with narrow rostrum (r) region at the apex (Table 1, developmental stage 4); Arron indicates sporophyte apex beneath calyptra. (B, C) Sporophyte with fully expanded sporangium and red annulus (a) (Table 1, developmental stage 9); (B) lines illustrate angle of sporangium inclination; (C) red peristome teeth (p) visible through the sporangium lid. (D) Air-dried sporophyte with a darker spore-filled region visible through the sporangium wall; lines illustrate measure of annulus diameter. (E) Annulus malformed and with a smaller diameter; the annulus is non-circular, penetrating the centre of the sporangium lid. (F) Peristome malformation with some peristome teeth fused to the inside of the sporangium lid; disabling sporangium dehiscence. (G) Sporangium lid naturally dehisced with lid now absent; revealing mature, properly formed, peristome teeth. Scale bars:(A, B, D) = 1 mm; (C, E, G) = 0·5 mm; (F) = 0·25 mm.(Budke, Goffinet and Jones, 2013)]

Tabela com os estágios de desenvolvimento da cápsula de Funaria spp.
Nas angiospermas, a cutícula tende a ser mais espessa no topo da folha (superfície adaxial), mas nem sempre é mais espessa. As folhas de plantas xerófitas adaptadas a climas mais secos têm espessuras de cutícula mais iguais em comparação com as de plantas de ambientes mesofíticos de climas mais úmidos, que não apresentam alto risco de desidratação por baixo das folhas.
"A lâmina cerosa da cutícula também funciona na defesa, formando uma barreira física que resiste à penetração de partículas de vírus, células bacterianas e os esporos e filamentos de hifas de fungos em crescimento.
As plantas terrestres precisam também se fixar ao substrato, para não serem levadas embora pela agua da chuva. Assim, as "briófitas", apresentam estruturas chamadas rizóides, com função de fixação.
Características gerais das
Hepáticas, Musci e Antóceros
Mesmo apresentando pequeno porte esses três clados juntos somam mais do que 18.000 espécies em todo o planeta.
As Hepáticas com cerca de 5.000 a 6.000 espécies, Musci ou Briofita com 10.000 a 13.000 espécies e Antocerófitas com 150 a 300 espécies.
Existem cerca de 9.500 espécies de musgos, sendo divididas em três classes: Andreaeidae (musgos-de-granito), Sphagnidae (musgos-de-turfeira) e Bryidae (conhecidos como “musgos verdadeiros”).
i) Metagênese
ii) Matrotrofia, a oosfera e o embrião retidos no gametófito femino
iii) Cloroplasto com tilacoide e grana
iv) Sem tecidos diferenciados
v) Corpo é um talo prostado, achatado ou folhoso
vi ) Tamanho pequeno
vii) Água e sais são transportados por difusão e osmose
viii) Geração gametofítica (haplóide) duradoura
ix) Geração esporofítica (diploide) efêmera
x) Esporófito se desenvolve como parasita do gametófito feminino
xi) Meiose espórica
xii) Uns apresentam poros aéreos, sem controle de abertura ou fechamento
xiii) Alguns apresentam estômatos (musgos) e Anthocerophyta possui estômatos no esporófito.
xiv) Apresentam cutícula com cutina
xv) Apresentam conceptáculos que produzem propágulos (gemas) que são levados pela água.
Simbiose "Briófitas"-Nostoc
Segundo Bustos-Díaz (2019) entre as "briófitas", as hepáticas Blasia e Calvicularia, junto com os Anthoceros, Phaeoceros, Notothylas e Dendroceros, estabelecem associações simbióticas bem definidas com cianobactérias (Meeks, 2005; Adams e Duggan, 2008).
A simbiose mais bem estudada desse grupo é Anthoceros punctatus-Nostoc. Essa simbiose é formada quando o Nostoc infecta a planta, que é atraída por seu hospedeiro após a liberação de HIFs (fatores indutores de hormogonia) no meio ambiente (hormogonia, ou células móveis). Uma vez que muitos hospedeiros produzem, ou acredita-se que produzam, fatores indutores de hormogonia (HIFs), o simbionte deve ter o equipamento genético necessário para produzir a célula móvel e detectar o gradiente de HIF que os guiará até o hospedeiro para formar a simbiose (Meeks e Elhai, 2002).
Após a infecção, o cianobionte se localiza nas cavidades viscosas presentes no talo gametófito. Quando a cianobacatéria Nostoc entra nesta região, a cavidade fecha-se para evitar mais infecções, criando as condições de baixo oxigênio necessárias para uma fixação aumentada de nitrogênio. Quando a simbiose é estabelecida, a quantidade de heterocistos aumenta acentuadamente, até 45% -50%, a atividade da nitrogenase aumenta e o fotossistema é desligado. A maior parte do nitrogênio fixado pelo cianobionte (80%) é transferido para o hospedeiro (Adams e Duggan, 2008), por meio de um mecanismo ainda não caracterizado.

MARCHANTIOPHYTAS
HEPÁTICAS
Grupo com 6.000 a 8.000 espécies.
Apresentam ciclo de vida gametofítico ou haploide dominante, com o esporófito dependente do gametófito, tanto fisicamente quando nutricionalmente.
Talo prostrado, achatado ou folhoso.
Os herboristas medievais ao descrever essa planta acharam que ela lembrava um fígado e colocaram o nome de hepática. O nome continuou o mesmo desde então. Hoje sabemos que é um grupo monofilético.

O ciclo de vida de M. polymorpha.
(A) Esquema do ciclo de vida de M. polymorpha
Diagrama de Prothallus baseado em Kny (1890). (B) Corte transversal do ápice do talo do tipo selvagem mostrando célula apical putativa (seta vermelha). (C) Talo selvagem mostrando ramificação dicotômica de entalhes apicais. (D) Anteridióforo do tipo selvagem. (E) arquegonióforo do tipo selvagem. (F) Anterídios do tipo selvagem. (G) arquegônia do tipo selvagem. (H) Esporófito jovem de tipo selvagem semelhante aos esporófitos sequenciados de 13 dias de Frank e Scanlon (2015). (I) Esporófitos maduros do tipo selvagem que se projetam de arquegonióforo. Asteriscos = entalhes apicais e gc = taças de gemas. (Flores-Sandoval e cols, 2018)

Blasia pusilla simbiose com Nostoc sphaericum e Nostoc punctiforme.
Porella navicularis e Nostoc sp.

Ciclo de vida de uma hepática (WP)

Ciclo de vida de Marchantia polymorpha L. 1753

Exemplos de musgos pleurocárpicos (A) e acrocárpicos (B-C). A: “tapete” do musgo pleurocárpico Pyrrhobryum spiniforme (Hook.) T. J. Kop. (Mniaceae), cobrindo um tronco de árvore. B: “tufos” de musgos acrocárpicos no solo. C: detalhe do esporófito na extremidade distal do gametófito do musgo acrocárpico Polytrichum juniperinum Willd. ex Hedw. (Polytrichaceae). Fotos: Tamara Matos (2019).(Matos, 2020).

Microscopia eletrônica de varredura da superfície do gametófito taloso de
Marchantia papillata Raddi (Marchantiaceae). As setas indicam o poro e a abertura
central para as trocas gasosas. B: conceptáculos (setas) na região dorsal do gametófito
taloso da hepática Marchantia sp. (Fotos A: Tamara Matos (2019); B: Déborah Santos, 2013). (Matos, 2020).

Gametófitos de Marchantia. A: detalhe da forma do gametófito masculino em forma de disco. B-C: gametófito feminino em forma de “guarda-chuva”. C: detalhe do gametófito feminino com os esporófitos (seta preta) e cápsulas (seta branca). (Imagens: Google Images. Matos, 2020)
ANTHOCEROPHYTA
Os Antóceros apresentam simbiose com a cianobactéria Nostoc spp, por isso apresentam uma cor verde escura profunda.

Estabelecem relação simbiótica com Nostoc spp
Arquegônio inserido no tecido do gametófito
Esporófito com columela onde são produzidos os esporos
Esporos em tétrades
Estômatos verdadeiros, abertura para ventilação e captura de CO2.
Os antóceros são um grupo monofilético, estabelecido já na idade média, com cinco famílias, 12 gêneros e 200 a 250 espécies no mundo; com hábitos terrícolas, saxícolas e epífitas. O gametófito dessas plantas, é taloso, pode abrigar colônias de cianobácterias (Nostoc sp), gerando uma importante simbiose. Por conta disse sua cor é de um verde profundo e escuro. Seu esporófito difere dos outros grupos de "briófitas" por crescer a partir de um meristema basal, que desenvolve esporos com germinação exospórica na maioria deles. O gênero Dendroceros conta com uma alta diversidade, por ter cerca de 40 espécies, das quais 13 estão presentes na região Neotrópical, presentes principalmente em regiões temperadas, subtemperadas e tropicais. Este gênero é o único exclusivamente epifítico, com esporos grandes, multicelulares e de germinação endospórica.

A. Dendroceros sp. B. Nothoceros sp. (setas indicam esporófitos) C. Notothylas sp. D. Anthoceros sp. (esporos e pseudoelatério em destaque) E. Phymatoceros sp. F. Phaeoceros sp. (esporos e pseudoelatério em destaque).

A – H. Dendroceros crispus (SW.) Nees. A. Esporo; B. Pseudoelatério; C. Gametófito; D. Gametófito com esporófito; E. Corte transversal do gametófito; F. Detalhe do corte transversal do gametófito; G. Células da parede da cápsula; H. Células da epiderme do gametófito com trigônios (espessamentos da parede). I – K. Nothoceros vincentianus (Lehm. & Lindenb.) Campb. I. Gametófito com esporófito; J. Esporo; K. Pseudoelatério. (A – H. Adaptado de Gradstein et al. 2001, Proskauer 1960; I – K. Adaptado de Gradstein et al. 2001).
MUSCI ou BRYOPHYTA
Diversidade
São registradas de 10.000 a 14.000 espécies em todas as zonas do planeta exceto na Antátdica
Corpo é um talo, sem órgãos e dividido em caulídio, filídio e rizoide
Estômatos unicelulares binucleados
Alguns chegam a 30 cm de altura como o Politrichum spp
Esporos germinam e originam protonema (semelhante a uma alga)
Protonema mecanismo de reprodução vegetativa pois se esplaha pelo solo
Apresentam Hidróides que se assemelham a xilema e Leptóide se assemelham a floema (sem feixes vasculares)

Reprodução dos musgos

Ciclo de vida de um musgo
VÍDEO BOTÂNICA
https://youtu.be/AqEXq6vt3r0

HEPÁTICA ou MARCHANTIOPHYTA
As plantas de Marchantia são morfologicamente simples. Eles crescem como um talo (tecido de folha plana que lembra uma lâmina de alga) com rizóides (filamentos semelhantes a raízes de uma única célula, com função de fixação) crescendo na superfície inferior e unidades repetitivas adaptadas para fotossíntese na superfície superior. Marchantia se reproduz sexualmente e assexuadamente. Propágulos assexuados (gemas) são formados dentro de estruturas especializadas (copos de gemas) que são dispersados por fatores mecânicos abióticos (por exemplo, gotas de chuva).

Gametófitos de Marchantia polymorpha L.
A esquerda o gametófito feminino (arquegonióforo) com gametóforos (arquegônios) a direita e o gamentófito masculino (anteridióforo) com seus gametóforos (anterídio).
(Foto: By Plantsurfer, in WP)

Marchantia polymorpha
Talo de Marchantia, propágulos.
Um talo maduro de Marchantia polymorpha mostrando
As As taças e as gemas em seu interior (propágulos). (Fonte: netnature)

Propágulos de Marchantia spp
Ao observar uma hepática procure na superfície superior por copos de gemas (taças) contendo gemas, pequenos discos verdes de células haplóides. Gemas são propágulos formados assexuadamente. Quando uma gota de chuva cai no copo de gemas (também chamado de “copo de respingo”), as gemas são espalhadas e dispersas para desenvolver gametófitos em outros lugares. Como os gemas são propágulos assexuados, os novos gametófitos serão geneticamente idênticos ao gametófito original, i.e., são clones.
(Foto Holger Casselmann)

Propágulos de Marchantia spp
(Foto de Franz Mattuschka)


Marchantiophyta
(Fonte: northerngardengems)

Marchantia polymorpha (Marchantiophyta)
(Fonte: netnature)


Arquegonióforo
Arquegonioforos de Marchantia . A. Primórdios dos Arquegonióforos. B. Arquegonióforos Jovem em desenvolvimento. C. Archaegoniophoro maduro (adulto) com células gaméticas prontas para serem coletadas (note a turbidez na gotícula de água).(Fonte: thenode) (no site thenode está ao contrário, acredito que houve um equívoco).

Anteridióforo
Anteridióforos de Marchantia spp. A. Primórdios dos Anteridióforos. B. anteridióforo jovem em desenvolvimento. C. Anteridióforos maduros. (Fonte: thenode).

Esporangio de Marchantia sp. A-C. Séries iniciais da esporogêneses.
D. Esporângio maduro. E. Detalhe do esporângio da foto D. F. Arquegonióforos fechados onde se desenvolveram arquegônios que depois da fecundação da oosfera formou o esporófito (no arquegônio). G. Esporófito com o esporângio aberto mostrando a liberação recente de esporos. H. Archegoniophoros com arquegônios (voltados para o solo) cujas oosfera já foram fecundadas. No esporófito há um tecido chamado esporângio que produz esporo por meiose. Detalhe dos esporângios abertos mostrando os esporos. (Fonte: thenode).

Arquegonióforo com arquegônios.
O arquegonióforo é a estrutura reprodutiva feminina que cresce a partir do talo. Identifique os arquegônios, que são em forma de vaso e pendem com suas aberturas tubulares voltadas para o solo. Cada arquegônio contém um único gameta feminino chamado oosfera (baruch).

O esporófito diplóide (2n) é cercado pelo arquegônio alargado chamado caliptra e é dependente do tecido haplóide do arquegonióforo para os nutrientes e água. O esporófito consiste em um pé onde se liga ao gametófito, uma haste chamada seta (ou cerda) e um grande esporângio em forma de ovo, no qual os esporos haploides (n) são produzidos por meiose (no esporângio).

Ciclo de vida de Marchantia polynorpha L.
Ciclo de vida de Marchantia polymorpha (Marchantia). Marchantia é uma hepática talóide que pode se reproduzir assexuadamente e sexualmente. Marchantia é uma planta dióica (i.e., os órgãos reprodutores masculinos e femininos se desenvolvem em talos distintos). O sexo é determinado pela presença de um cromossomo sexual Y no caso das plantas macho e X no caso das plantas fêmeas. A fase dominante haplóide é constituída principalmente por uma estrutura semelhante a uma folha (bidimensional paralela ao subistrato) denominada talo (gametófito) (1). Propágulos assexuados conhecidos como gemmae (singular: gemma) (2) são produzidos dentro de uma estrutura especializada chamada copo ou taça de gemas (gemma cup) (3) que se desenvolve no lado dorsal do talo. As gemas multicelulares desenvolvem-se em talo, completando assim o ciclo assexuado (4).
As gemas contêm células-tronco apicais meristemáticas (do meristema apical) (também conhecidas como incisuras ou entalhes apicais) localizadas nas incisuras (fendas) nas extremidades do talo que direcionam o crescimento do talo em um padrão bifurcado característico desta espécie. Quando os talos maduros são expostos a uma combinação particular de luz vermelha distante ou vermelho extremo (far-red) (um pouco antes do infra vermelho) e luz branca, arquegonióforos (5) abrigando arquegônia (6) (singular: arquegônio) e anteridióforos (7) abrigando anterídia (8) (singular: anterídium) são formados diretamente a partir das incisuras (fendas) apicais dos talos maduros.
Os espermatozóides (anterozóides) móveis (9) produzidos nos anterídios aproveitam as gotas de água (10) (a chuva é a fonte mais comum de água) para nadar em direção ao arquegônio (11) onde fertilizam os óvulos (oosfera) (12). Há uma oosfera em cada arquegônio. O embrião (13) se desenvolve dentro de uma caliptra (14) que protege e nutre o esporófito em crescimento, quando o esporófito esta maduro e após a meiose do esporângio este dará origem aos esporos (15). Milhares de esporos contidos nos esporângios do esporófito (16) são liberados (17) e se desenvolvem em esporos jovens que ao germinarem formarão o protonema que dará origem a um novo talo adulto maduro, completando assim o ciclo sexual.(modif. de Aguilar-Cruz, 2019).
Vermelho distante ou Far-red

A luz vermelho extremo ou luz vermelho distante é uma faixa de luz na extremidade vermelha do espectro visível, logo antes da luz infravermelha. Geralmente considerada como a região entre 700 e 780 nm, é fracamente visível aos olhos humanos. É amplamente refletida ou transmitida pelas plantas devido ao espectro de absorbância da clorofila e é percebida pelo fitocromo fotorreceptor da planta. No entanto, alguns organismos podem usá-lo como fonte de energia na fotossíntese. A luz vermelha distante também é usada para a visão por certos organismos, como algumas espécies de peixes de grande profundidade (WP).
VÍDEO

Cápsula de um musgo



Em destaque Marchantiophyta, Anthocerophyta e Musci ou Bryophyta, as três primeiras divisões que invadiram o ambiente terrestre úmido.

Plantas terrestres

Viridiplantae


Hipótese de relacionamento entre os diferentes grupos de Archaeplastida.
A oogamia é uma forma de anisogamia (heterogamia) na qual o gameta feminino é significativamente maior que o gameta masculino, sendo também imóvel. Os gametas masculinos são tipicamente espermatozóides com grande mobilidade, competindo pela fertilização do óvulo. A oogamia é predominante nos animais, mas também pode ocorrer em protistas, algumas ordens de algas e em alguns vegetais como briófitas, fetos, e algumas gimnospérmicas como as cicadáceas e Ginkgo. Em algumas algas, na maioria das gimnospermas e nas angiospermas, uma variante de oogamia ocorre onde as células reprodutoras masculinas não são móveis (WP).

Cladograma que apresenta alguns dos caracteres compartilhados pelas algas verdes e as plantas terrestres (Raven et al, 1992).
https://www.semanticscholar.org/paper/Establishment-of-a-functional-symbiosis-between-the-Wong-Meeks/900bb35d9668810f6fd299274efb91b9c45008cc
https://www.sciencedirect.com/topics/agricultural-and-biological-sciences/anthoceros
https://nph.onlinelibrary.wiley.com/doi/full/10.1111/nph.15191
https://netnature.wordpress.com/2014/05/28/botanica-de-briofitas-bryopsida-marchantiophyta-anthocerotophyta/
http://azolla.fc.ul.pt/aulas/documents/BotAulas9_10_11_12.pdf
https://www.quora.com/What-are-reduced-gametophytes-What-are-the-advantages-of-them-in-seed-plants
https://tophat.com/marketplace/science-&math/biology/textbooks/openstax-biology-concepts-openstax-content/80/4192/
https://tophat.com/marketplace/science-&-math/biology/textbooks/openstax-biology-concepts-openstax-content/80/4192/
https://pt.wikipedia.org/wiki/Embryophyta
http://www.plantcell.org/content/31/9/1934
https://www.scielo.br/pdf/cr/v35n3/a43v35n3.pdf
http://www.fisiologiavegetal.ufc.br/Aulas%20em%20PDF%20PG/Unidade%20VI1.pdf
https://en.wikipedia.org/wiki/Archaeplastida
https://www.researchgate.net/publication/297301899_Microrganismos_x_Planta_guerra_ou_parceria
https://www.sciencedirect.com/science/article/pii/S0960982213005046
https://edisciplinas.usp.br/pluginfile.php/5588303/mod_resource/content/1/BIB304_caract%20grandes%20grupos_2020.pdf
https://reader.elsevier.com/reader/sd/pii/S0960982213005046?token=EA3D99686B21BCD5D6EA8D310530C0D998DC9CD58658D2C5373C35CDFC4EC0026E747B97CF18A9CC9AE654E0227C1031&originRegion=us-east-1&originCreation=20210515150118
http://botanicaonline.com.br/geral/arquivos/Texto%20Base%20Aula%2011.pdf
https://pt.wikipedia.org/wiki/Oogamia
https://www.sciencedirect.com/topics/agricultural-and-biological-sciences/anthoceros
https://nph.onlinelibrary.wiley.com/doi/full/10.1111/nph.15191
https://netnature.wordpress.com/2014/05/28/botanica-de-briofitas-bryopsida-marchantiophyta-anthocerotophyta/
http://azolla.fc.ul.pt/aulas/documents/BotAulas9_10_11_12.pdf
https://www.quora.com/What-are-reduced-gametophytes-What-are-the-advantages-of-them-in-seed-plants
https://tophat.com/marketplace/science-&math/biology/textbooks/openstax-biology-concepts-openstax-content/80/4192/
https://tophat.com/marketplace/science-&-math/biology/textbooks/openstax-biology-concepts-openstax-content/80/4192/
https://pt.wikipedia.org/wiki/Embryophyta
http://www.plantcell.org/content/31/9/1934
https://www.scielo.br/pdf/cr/v35n3/a43v35n3.pdf
http://www.fisiologiavegetal.ufc.br/Aulas%20em%20PDF%20PG/Unidade%20VI1.pdf
https://en.wikipedia.org/wiki/Archaeplastida
https://www.researchgate.net/publication/297301899_Microrganismos_x_Planta_guerra_ou_parceria
https://www.sciencedirect.com/science/article/pii/S0960982213005046
https://edisciplinas.usp.br/pluginfile.php/5588303/mod_resource/content/1/BIB304_caract%20grandes%20grupos_2020.pdf
https://reader.elsevier.com/reader/sd/pii/S0960982213005046?token=EA3D99686B21BCD5D6EA8D310530C0D998DC9CD58658D2C5373C35CDFC4EC0026E747B97CF18A9CC9AE654E0227C1031&originRegion=us-east-1&originCreation=20210515150118
http://botanicaonline.com.br/geral/arquivos/Texto%20Base%20Aula%2011.pdf
https://pt.wikipedia.org/wiki/Oogamia
http://www.esalq.usp.br/lepse/imgs/conteudo_thumb/Evolu--o-das-plantas-o-come-o-de-tudo--parte-1.pdf
https://teses.usp.br/teses/disponiveis/41/41132/tde-22062020-091350/publico/Tamara_Matos_CORRIG.pdf
Protistas
https://rhinellalab.com/protistas
https://en.wikipedia.org/wiki/Pectin
ESCÓLIOS
https://en.wikipedia.org/wiki/Pectin
(Publicado em 16/IV/2020 e atualizado em 15/IV/2022)
ESCÓLIOS
Sagan, L. (1967). On the origin of mitosing cells. J. Theor. Biol.14, 255-74
A theory of the origin of eukaryotic cells (“higher” cells which divide by classical mitosis) is presented. By hypothesis, three fundamental organelles the mitochondria, the photosynthetic plastids and the (9+2) basal bodies of flagella were themselves once free-living (prokaryotic) cells. The evolution of photosynthesis under the anaerobic conditions of the early atmosphere to form anaerobic bacteria, photosynthetic bacteria and eventually blue-green algae (and protoplastids) is described. The subsequent evolution of aerobic metabolism in prokaryotes to form aerobic bacteria (protoflagella and protomitochondria) presumably occurred during the transition to the oxidizing atmosphere. Classical mitosis evolved in protozoan-type cells millions of years after the evolution of photosynthesis. A plausible scheme for the origin of classical mitosis in primitive amoeboflagellates is presented. During the course of the evolution of mitosis, photosynthetic plastids (themselves derived from prokaryotes) were symbiotically acquired by some of these protozoans to form the eukaryotic algae and the green plants. The cytological, biochemical and paleontological evidence for this theory is presented, along with suggestions for further possible experimental verification. The implications of this scheme for the systematics of the lower organisms is discussed.
Symbiogenesis is an evolutionary mechanism caused by hereditary symbiosis. Symbiogenesis theories were introduced in early twentieth-century Russia, when Merezhkowsky, Faminstyn, and Kozo-Polyansky recognized that organellar structures present in the eukaryotic cell evolved through symbiogenesis. Symbiogenesis research subsequently spread to Europe and the Americas through the works of von Faber, Portier, Buchner, Schneider, Wallin, and Lederberg. Nonetheless, symbiogenesis theories were excluded from the Modern Synthesis that advanced a selectionist account of evolution, and were only reintroduced by Margulis, from the 1960s onward. Her Serial Endosymbiotic Theory gives a synthetic account of how symbiogenesis underlies the origin of the four eukaryotic kingdoms. Today, symbiogenesis research associates with research on lateral gene transfer, mobile genetic elements, viriome and microbiome studies, and the Gaia Hypothesis, making symbiogenesis one of the candidates for an Extended Synthesis.
Current Biology23, 1081–1084, June 17, 2013
Shinichiro Maruyama and Eunsoo Kim
Green algae, land plants, and other photosynthetic eukary-otes possess plastids, such as chloroplasts, which haveevolved from cyanobacterial ancestors via endosymbiosis[1]. An early evolutionary merger between heterotrophic eu-karyotes and cyanobacteria called primary endosymbiosisgave rise to the first photosynthetic eukaryotes. A seriesof plastid acquisitions involving engulfment of eukaryoticphototrophs, known as secondary or tertiary endosymbi-osis, followed [2]. Through these repeated symbiotic events,photosynthesis spread across a number of eukaryotic line-ages [2, 3 ]. While the origin of eukaryotic photosynthesiswas undoubtedly a fundamentally important evolutionaryevent in Earth’s history, without which much of the modernmarine phytoplankton would not exist, the cellular pro-cesses that shaped this initial plastid genesis remain largelyunknown. Here, we report ultrastructural evidence for bacte-rial phagocytosis in a primary plastid-bearing alga. This mix-otrophic green alga utilizes a mouth-like opening, a tubularchannel, and a large permanent vacuole to engulf, transport,and digest bacterial cells. This mode of phagocytosis, likelyinherited from its plastid-lacking ancestor, differs fromthose displayed by many other eukaryotes, including ani-mals, amoebas, and ciliates. These results provide insightinto the key phagocytosis step during the origin of the firstphotosynthetic eukaryotes.
Ann Bot. 2013 May; 111(5): 781–789.
As the ancestrally dominant generation that both protected and nourished the zygote, the gametophyte played a significant role in the evolution of the multicellular sporophyte in embryophytes. Subsequent reductions in the gametophyte have decreased its influence in the vast majority of plants. However, in bryophytes the gametophyte generation remains influential; sporophyte offspring are in contact with, nourished from, and partially surrounded by the maternal gametophyte throughout their lifespan (Ligrone et al., 1993; Graham and Wilcox, 2000). This prolonged interaction may result in maternal effects that play a significant role in the survival, development, and fitness of the offspring sporophytes.
Protection against the desiccative effects of life above water is critical for the survival of terrestrial plants. An array of adaptations has evolved that decrease water loss from plant bodies (e.g. cuticle, bark, trichomes, scales; Esau, 1977). Of these structures, the plant cuticle is ubiquitous, occurring in taxa from all major lineages of land plants [e.g. mosses (Proctor, 1979), cheilanthoid ferns (Sigel et al., 2011), gymnosperms, Ginkgo (Taylor et al., 1989) and angiosperms, Agave (Wattendorff and Holloway, 1980)]. The cuticle consists of layers of the biopolymers cutin/cutan and waxes that cover and permeate the outermost layers of the cell wall (Holloway, 1982), decreasing intracellular water loss. Cuticle function relating to water loss has been studied primarily in angiosperms and less so in other tracheophyte lineages (e.g. Riederer and Schreiber, 2001; Schreiber and Schönherr, 2009). Cuticles on the leafy gametophytes of mosses may also function in decreasing water loss (i.e. endohydric taxa; Proctor, 1979). Herein we demonstrate experimentally that the cuticle on the maternal moss gametophyte plays a critical role in reducing dehydration stress on developing sporophyte offspring.
The moss sporophyte apex is covered throughout much of its development by the calyptra, a cap of maternal gametophytic tissue. For decades the calyptra has been known to be critical for both sporangium differentiation and sporogenesis (Herzfelder, 1923; Bopp, 1954; French and Paolillo, 1975). The calyptra has been hypothesized to influence the sporophyte in various ways including, mechanical constraint (physical), hormonal secretion (physiological), and desiccation protection. In terms of desiccation protection, we know that young sporophytes survive removal of the calyptrae only when grown at high humidities (Bopp, 1957; French and Paolillo, 1975) and a dye solution ascends sporophytes faster without calyptrae compared with those with calyptrae on the apices (Bopp and Stehle, 1957), suggesting a greater rate of water loss from sporophytes lacking a calyptra. A role in controlling internal sporophyte water balance suggests that the calyptra is covered by a cuticle, a hypothesis raised as early as 1884 by Hy, and only recently confirmed (Hy, 1884; Budke et al., 2011). Determining whether the calyptra cuticle contributes to dehydration protection of the developing sporophyte may add further support to the calyptra's role in maternal protection.
In Funaria hygrometrica the calyptra is covered by a multilayered cuticle that is significantly thicker than that covering the sporophyte or leafy gametophyte, which is fully developed prior to elongation of the underlying sporophyte (Budke et al., 2011). Furthermore, the development of a cuticle on the sporophyte apex is completed only during the maturation of the sporogenous capsule (Budke et al., 2012). Thus water loss from the developing sporophyte is likely to be prevented by the maternal calyptra rather than by structures of the sporophyte itself. While the significance of the calyptra to sporophyte maturation has been demonstrated, the effects on different components of the maturation process remain obscure.
Despite its small size, the calyptra and its cuticle are reasonably amenable to experimentation. The calyptra can be physically removed from the sporophyte, the cuticle waxes chemically extracted, and then replaced on the sporophyte apex. Calyptra vitality is most likely unnecessary for later stages of sporophyte development (Bopp, 1957; French and Paolillo, 1975), and thus any cellular damage caused by the chemical removal of the cuticle is experimentally acceptable. To examine the importance of the maternal calyptra cuticle for the sporophyte offspring, we experimentally removed the calyptra cuticle of the moss F. hygrometrica and then exposed the sporophytes to a short-term dehydration event. If the function of the calyptra cuticle is to prevent dehydration, then we predict that components of sporophyte fitness (i.e. survival, development, functional morphology, reproductive output) will be negatively affected when the calyptra cuticle has been removed compared with those with the calyptra cuticle intact.(Budke, Goffinet and Jones, 2013)
CAROTENOIDES
Os carotenóides compreendem uma família de compostos naturais, dos quais mais de 600 variantes estruturais estão reportadas e caracterizadas, a partir de bactérias, algas, fungos e plantas superiores. A produção natural mundial é estimada em 100 milhões de toneladas por ano, e é encabeçada pela fucoxantina das algas fotossintéticas marrons. Os carotenóides mais comumente encontrados nos vegetais são: o b-caroteno (cenoura; Daucus carota), licopeno (tomate; Lycopersicum esculentum), várias xantofilas (zeaxantina, luteína e outras estruturas oxigenadas do milho, Zea mays; da manga, Mango indica; do mamão, Carica papaya) e a bixina (aditivo culinário e corante dérmico usado por indígenas amazônicos, obtido do urucum, Bixa orellana) (BRITTON et al.,1995).
Os carotenóides são tetraterpenóides de 40 carbonos unidos por unidades opostas no centro da molécula (FRASER & BRAMLEY,2004). Ciclização, hidrogenação, desidrogenação, migração de duplas ligações, encurtamento ou alongamento da cadeia, rearranjo, isomerização, introdução de funções com oxigênio ou a combinação destes processos resultam na diversidade de estruturas dos carotenóides (DELGADO-VARGAS, 2000).
Todos os diferentes tipos de carotenóides encontrados nos organismos fotossintéticos são moléculas lineares com múltiplas cadeias duplas conjugadas (Figura 2). A cadeia poliênica pode ter de 3 a 15 duplas ligações conjugadas e o comprimento do cromóforo determina o espectro de absorção e a cor da molécula. Todas são baseadas em 7 diferentes grupos terminais, dos quais somente 4 (β, ε, κ e ψ) são encontradas em carotenóides de vegetais superiores (FRASER & BRAMLEY, 2004). Os carotenóides compostos somente de carbono e hidrogênio são chamados de carotenos e os carotenóides oxidados, as xantofilas, apresentam grupos substituintes com oxigênio, como hidroxilas, grupos ceto e epóxi (DELGADO-VARGAS, 2000; OLIVIER & PALOU, 2000).
Os carotenóides normalmente apresentam duas a três bandas de absorção localizadas na região do azul-violeta do espectro. Por exemplo, o β-caroteno dissolvido em hexano, apresenta uma banda de absorção na faixa de 430, 450 e 480 nm (GOVINDJEE, 1969).

Tabela. Características de absorção de alguns carotenóides (Fonte: GOVINDJEE, 1969).
0 comentários:
Postar um comentário