DOMÍNIO ARCHAEA
Reino Archaea

Lado esquerdo: Methanosarcina, Ignicoccus com Nanoarchaeum, ARMAN e Haloquadratum. Na direita: Methanohalophilus, Pyrococcus, Prometheoarchaeum e Halobacterium.
(Fonte: Maulucioni, in WP)

Lago Magadi, Kenia.
As haloarqueas que crescen no lago Magadi no Kenia são de cores púrpuras e avermelhadas devido a pigmentos carotenoides como la bacterioruberina.
O termo ‘Archaea’ é derivado de palavra grega, ἀρχαῖος, archaĩos, que significa primitivo ou antigo, usada para indicar as caracaterísticas "ancestrais" desses organismos quando comparadas com Eukarya, não que estejam na base da árvore filogenética de todos os organismos vivos. Como veremos eles estão mais proximos aos Eukarya devido as suas características.
Archaea e bactérias são geralmente semelhantes em tamanho e forma, embora algumas archaea tenham formas muito diferentes, como as células planas e quadradas de Haloquadratum walsbyi.
Apesar da semelhança morfológica com as bactérias, as archaea possuem genes e várias vias metabólicas que estão mais intimamente relacionadas às dos eucariotos, principalmente pelas enzimas envolvidas na transcrição e tradução.
Outros aspectos da bioquímica archaeal são únicos, como sua dependência de éter lipídios em suas membranas celulares, incluindo archaeois.
Archaea usam fontes de energia mais diversas do que eucariotos, variando de compostos orgânicos, como açúcares, amônia, íons metálicos ou até gás hidrogênio.
A Haloarchaea tolerante ao sal usa a luz solar como fonte de energia, e outras espécies de archaea fixam carbono, mas ao contrário de plantas e cianobactérias, nenhuma espécie conhecida de archaea faz as duas coisas.
Archaea se reproduzem assexuadamente por fissão binária, fragmentação ou brotamento; ao contrário das bactérias, nenhuma espécie conhecida de Archaea forma endósporos.
As primeiras arqueias observadas eram extremófilos, vivendo em ambientes extremos, como fontes termais e lagos salgados, sem outros organismos. Ferramentas aprimoradas de detecção molecular levaram à descoberta de archaea em quase todos os habitats, incluindo solo, oceanos e pântanos. Archaea são particularmente numerosas nos oceanos, e as archaea no plâncton marinho podem ser um dos grupos de organismos mais abundantes em nosso planeta.
Archaea são uma parte importante da vida da Terra. Eles fazem parte da microbiota de todos os organismos. No microbioma humano, eles são importantes no intestino, na boca e na pele.
Sua diversidade morfológica, metabólica e geográfica permite que desempenhem múltiplos papéis ecológicos: fixação de carbono; ciclagem de nitrogênio; e manutenção de comunidades microbianas simbióticas e sintróficas.
Não são conhecidos exemplos claros de arquéias patogênicos ou parasitas. Em vez disso, muitas vezes são mutualistas ou comensais, como os metanogênicos (cepas produtoras de metano) que habitam o trato gastrointestinal em humanos e ruminantes, onde seu grande número facilita a digestão. Metanogênicos também são usados na produção de biogás e tratamento de esgoto, e a biotecnologia explora enzimas de arqueas extremófilas que podem suportar altas temperaturas e solventes orgânicos.
História
Archaea, naquela época apenas os metanogênicos eram conhecidos, foram classificados pela primeira vez separadamente das bactérias em 1977 por Carl Woese e George E. Fox com base na comparação da sequência dos genes de RNA ribossômico (rRNA).
Eles chamaram esses grupos de UR-Reinos Archaebacteria e Eubacteria, embora outros pesquisadores os tratassem como reinos ou sub-reinos.
Woese e Fox deram a primeira evidência para Archaebacteria como uma "linha de descendência" separada:
1. falta de peptidoglicano em suas paredes celulares,
2. duas coenzimas incomuns,
3. resultados do sequenciamento do gene RNA ribossômico 16S.
Para enfatizar essa diferença, Carl R. Woese, Otto Kandler e Mark Wheelis propuseram mais tarde a reclassificação dos organismos em três domínios naturais conhecidos como o sistema de três domínios: o Archaea, Bacteria e Eukarya no que agora alguns historiadores da ciência chamam de Revolução Woesiana.
A palavra ARCHAEA como vimos vem do grego antigo ἀρχαῖα, que significa "coisas antigas", uma vez que os primeiros representantes do domínio Archaea eram metanógenos e extremófilos e supunha-se que seu metabolismo refletia a atmosfera primitiva da Terra e a antiguidade dos organismos, mas como novos habitats foram estudados, mais organismos foram descobertos.
Arquéas halofílicos extremos e hipertermofílicos também foram incluídos em Archaea. Por muito tempo, as arquéas foram vistas como extremófilos que existem apenas em habitats extremos, como fontes termais e lagos salgados, entretanto no final do século XX, as arquéas também foram identificadas em ambientes não extremos, como o ambiente marinho, o solo e nossa pele.
Hoje, eles são conhecidos por serem um grupo grande e diversificado de organismos abundantemente distribuídos por todos os ambientes.
A constatação da importância e da onipresença das arquéa veio do uso da reação em cadeia da polimerase (PCR) para detectar procariotos de amostras ambientais (como água ou solo) multiplicando seus genes ribossômicos. Isso permite a detecção e identificação de organismos que não foram cultivados em laboratório.
Um domínio antigo e misterioso
Desde a proposta de um domínio Archaea em 1977, estudos revelam que os organismos arquéa possuem características semelhantes tanto a procariotos como eucariotos, mas também possuem características próprias suas ou sinapomorfias.
Dentre suas características distintivas, temos a composição da membrana citoplasmática, que é formada por cadeias de isopreno ligadas às moléculas de glicerol-1-fosfato por meio de ligações do tipo éter, enquanto bactérias e eucariotos possuem cadeias de ácidos graxos que se ligam ao glicerol-3-fosfato por uma ligação do tipo éster (Kates et al., 1993).
Algumas membranas de Archaea organizam-se como monocamadas lipídicas, ao invés da tradicional bicamada encontrada nos outros dois domínios (Bullock, 2000). Este tipo de organização é geralmente encontrado em arquéas hipertermófilas, conferindo maior fluidez à membrana desses organismos (DeRosa et al., 1991).
Outra característica peculiar deste domínio consiste na ausência de peptideoglicano em suas paredes celulares, propriedade comum a todas as bactérias. Ao invés disso, as archaeas possuem uma grande variedade de envoltórios celulares, como pseudopeptideoglicano, metanocondroitina, heteropolissacarídeos sulfatados, glutaminilglicano e membrana externa (Kandler & König, 1998; Albers & Meyer, 2011).
Pode existir ainda, associada a outros envoltórios celulares ou como único envoltório, uma camada de proteínas ou glicoproteínas que formam arranjos cristalinos simétricos e com capacidade de auto-organização, denominadas camada S (Konig et al, 2007).
Estruturas de superfície de archaeas, como fímbrias e flagelos também apresentam composições diferentes das estruturas de bactérias (Ng et al., 2008).
Em relação aos processos celulares, a maquinaria de processamento de informação, como divisão celular, estrutura do DNA, citoesqueleto, transcrição e tradução de archaeas se assemelham mais à maquinaria de células eucarióticas, podendo ser considerada uma versão simplificada desta (Ishino & Ishino, 2012).
A RNA polimerase de Archaea possui estrutura e função semelhantes à RNA polimerase II de eucariotos (Zillig et al., 1979; Stetter et al., 1980), sendo observada nelas, uma resistência a antibióticos que afetam as RNA polimerases de bactérias (Sturm et al., 1980).
A RNApolimerase, depende de um menor número de fatores de transcrição (Grohmann & Werner, 2011). Os ribossomos de Archaea apresentam coeficiente de sedimentação 70S, como as bactérias, porém com uma composição proteica diferente.
O número de fatores traducionais em Archaea é muito maior do que o observado em Bacteria e apresentam um alto grau de homologia com os fatores eucarióticos (Dennis, 1997). Por outro lado, os genes operacionais, envolvidos no metabolismo energético e vias biossintéticas se assemelham mais aos genes bacterianos (Spang et al., 2013).
O primeiro grupo de arqueas estudado foi o das "bactérias metanogênicas ou metanógenas".
A metanogênese foi descoberta no lago Maior de Itália em 1776, ao observar nele o borbulhar do "ar combustível". Em 1882 observou-se que a produção de metano no intestino dos animais devia-se à presença de micro-organismos (Popoff, Tappeiner e Hoppe-Seyler).
Em 1936, ano que marcou o princípio da era moderna no estudo da metanogênese, H.A Barker oferece as bases científicas para o estudo da sua fisiologia e conseguiu desenvolver um meio de cultivo apropriado para o crescimento dos metanógenos. Nesse ano foram identificados os géneros Methanococcus e Methanosarcina.
As primeiras arqueias extremófilas foram encontradas em ambientes quentes. Em 1970, Thomas D. Brock, da Universidade de Wisconsin, descobriu a Thermoplasma, uma arqueia termoacidófila, e em 1972 a Sulfolobus, uma hipertermófila.
Brock se iniciou em 1969 no campo da biologia dos hipertermófilos com a descoberta de Thermus aquaticus, que não é uma arquea mas antes uma bactéria.
Em 1977 identificam-se as arqueias como o grupo procarionte mais distante ao descobrir que os metanógenos apresentam uma profunda divergência com todas as bactérias estudadas. Nesse mesmo ano propôs-se a categoria de super-reino para este grupo com o nome de Archaebacteria. Atualmente as Archaea estão no seu próprio reino, o reino Archaea em um domínio próprio.

Os três domínios dos organismos vivos. Bactérias e Archaea são procariontes, mas diferem o suficiente para serem colocadas em domínios separados. Acredita-se que um ancestral das modernas Archaea deu origem ao Eukarya, o terceiro domínio da vida. Alguns grupos de Archaea e Bacteria são mostrados.
As arquéas, que há algum tempo atrás eram chamadas de arqueobactérias, e estavam colocadas juntos com as Eubactérias (num mesmo resino), são organismos procariontes pertencentes ao domínio Archaea.
Elas apresentam um conjunto de características que justifica serem incluídas em um reino distinto do reino das Eubactérias, diferenciando-se destas por:
1) mRNA
O RNA desses organismos assemelha-se mais ao dos eucariotes do que das bactérias.
2) tRNA
As moléculas de tRNA são únicas e compartilham muitas características comuns com os tRNA eucariotos do que com os tRNA dos procariotos.
3) DNA
A replicação do DNA é um processo celular fundamental que é funcionalmente conservado em todos os três domínios da vida (Bacteria, Archaea e Eukarya). A regulação precisa da replicação do DNA garante a duplicação precisa da informação genômica, e a iniciação da replicação é o primeiro e mais importante estágio dessa regulação. O primeiro modelo de iniciação da replicação do DNA foi proposto para Escherichia coli em 1963, postulando que um fator de ação trans se liga a um sítio de ação cis que desencadeia a iniciação da replicação (Jacob et al., 1963). Nos 50 anos subsequentes, esse “modelo de replicon” demonstrou ser essencialmente verdadeiro em todos os organismos, e o sítio de ação cis agora é conhecido como origem de replicação.

Distribuição das origens de replicação do DNA em três sistemas modelo arqueais bem estudados, espécies de Pyrococcus (A), espécies de Sulfolobus (B) e Haloarcula hispanica (C). As origens são indicadas com ovais preenchidos e as pontas de seta designam a replicação bidirecional. Replicador-iniciador indica que cada origem é especificada por seu iniciador codificado proximalmente. Tanto as espécies de Pyrococcus quanto as de Sulfolobus contêm um único cromossomo; o cromossomo das espécies de Pyrococcus é replicado a partir de uma única origem (Myllykallio et al., 2000), enquanto o cromossomo das espécies de Sulfolobus é replicado de três origens em quase sincronia (Duggin et al., 2008). O genoma de Haloarcula hispanica consiste em um cromossomo principal e dois elementos extracromossômicos com cinco origens de replicação ativas: oriC1-cdc6A e oriC2-cdc6E no cromossomo I principal, oriC6-cdc6I e oriC7-cdc6J no minicromossomo II e oriP-cdc6K no megaplasmídeo pHH400 (Wu et al., 2012).
Conclusão: Replicação no domínio Archaea, inicia em mais de um ponto com nos cromossomos dos eucariotos.
Dominio Bacteria.
Os cromossomos bacterianos são tipicamente replicados a partir de uma única origem.
Domínio Eukarya.
A replicação dos cromossomos eucarióticos inicia a partir de várias origens discretas (Leonard e Mechali, 2013). As origens de replicação do DNA foram bem definidas em bactérias e eucariotos unicelulares.
Dominio Archaea. Em contraste, o foco nas origens de replicação do DNA em archaea, o terceiro domínio da vida, começou apenas aproximadamente uma década atrás. As origens de replicação do DNA foram mapeadas principalmente para alguns representantes de espécies de arqueias distribuídas nos três principais filos, Euryarchaeota, Crenarchaeota e Thaumarchaeota (Myllykallio et al., 2000; Lundgren et al., 2004; Robinson et al., 2004; Grainge et al., 2006; Norais et al., 2007; Majernik e Chong, 2008; Coker et al., 2009; Pelve et al., 2012, 2013; Wu et al., 2012, 2014).
Além disso, caracterização mais detalhada foi revelada em vários sistemas modelo, como espécies de Pyrococcus (Myllykallio et al., 2000; Matsunaga et al., 2001, 2003), espécies de Sulfolobus (Lundgren et al., 2004; Robinson et al. , 2004; Duggin et al., 2008; Samson et al., 2013), Haloferax volcanii (Norais et al., 2007; Hawkins et al., 2013) e Haloarcula hispanica (Wu et al., 2012, 2014).
Sabe-se agora que as archaea usam uma ou várias origens para replicar seus cromossomos circulares (Kelman e Kelman, 2004; Robinson e Bell, 2005; Hyrien et al., 2013). As múltiplas origens, juntamente com seus genes iniciadores adjacentes em certos cromossomos archaeais, podem ter surgido da captura de elementos extracromossômicos e parecem ser mosaicos de pares replicadores-iniciadores distintos (Robinson e Bell, 2007; Wu et al., 2012).
O DNA das arquéas apresenta introns interrompendo os genes em seu cromossomo, característica esta, presente também em todos os Eukarya.
4) Membrana plasmática
A membrana plasmática procariótica é uma bicamada lipídica fina com 6 a 8 nn, nanômetros) que envolve completamente a célula e separa o interior do exterior. Sua natureza seletivamente permeável mantém íons, proteínas e outras moléculas dentro da célula e impede que eles se difundam no ambiente extracelular, enquanto outras moléculas podem se mover através da membrana. Lembre-se de que a estrutura geral de uma membrana celular é uma bicamada fosfolipídica (composta de duas camadas de moléculas lipídicas). Nas membranas das células das arqueias as cadeias de isopreno (fitanil) ligadas ao glicerol substituem os ácidos graxos ligados ao glicerol nas membranas bacterianas. Algumas membranas plasmátaicas são monocamadas de lipídios em vez de bicamadas.

Os fosfolipídeos das arquéias diferem daqueles encontrados em Bactéria e Eukarya de duas maneiras. Primeiro, eles têm cadeias laterais de fitanil ramificadas em vez de lineares. Em segundo lugar, uma ligação de éter em vez de uma ligação de éster conecta o lipídio ao glicerol. (Courses).
Resumindo: a membrana plasmática das arquéas apresenta lipídeos compostos de uma associação de glicerol-éter, enquanto que os das bactérias e eucariotos são compostos de glicerol-éster.
3) A parede celular
O citoplasma das células procarióticas tem uma alta concentração de solutos dissolvidos. Portanto, a pressão osmótica dentro da célula é relativamente alta. A parede celular é uma camada protetora que envolve as células e lhes dá forma e rigidez. Ele está localizada por fora da membrana celular e evita a lise osmótica (rompimento devido ao aumento do volume). A composição química da parede celular varia entre espécies de Bacteria e Archaea.
As paredes das células bacterianas contêm peptidioglicano, composto de cadeias polissacarídicas que são reticuladas por peptídeos incomuns contendo aminoácidos L e D, incluindo ácido D-glutâmico e D-alanina. (As proteínas normalmente têm apenas L-aminoácidos; como consequência, muitos de nossos antibióticos atuam imitando D-aminoácidos e, portanto, têm efeitos específicos no desenvolvimento da parede celular bacteriana.)
Existem mais de 100 formas diferentes de peptidioglicano. As proteínas da camada S (camada superficial) também estão presentes na parte externa das paredes celulares de Archaea e Bacteria.
As bactérias são divididas em dois grupos principais: Gram positivas e Gram negativas, com base em sua reação à coloração de Gram. Observe que todas as bactérias Gram-positivas pertencem a um filo; bactérias em outros filos (Proteobacteria, Chlamydias, Spirochetes, Cyanobacteria e outras) são Gram-negativas.
O método de coloração de Gram recebeu o nome de seu inventor, o cientista dinamarquês Hans Christian Gram (1853–1938). As diferentes respostas bacterianas ao procedimento de coloração são, em última análise, devido à estrutura da parede celular (e a sua composição). Os organismos Gram-positivos normalmente não têm a membrana externa (a cápsula bacteriana) encontrada nos organismos Gram-negativos. Até 90 por cento da parede celular em bactérias Gram-positivas é composta de peptidoglicano, e a maior parte do resto é composta de substâncias ácidas chamadas ácidos teicóicos. Os ácidos teicóicos podem ser covalentemente ligados a lipídios na membrana plasmática para formar ácidos lipoteicóicos. Os ácidos lipoteicóicos ancoram a parede celular à membrana plasmática.
As bactérias Gram-negativas têm uma parede celular relativamente fina composta por algumas camadas de peptidoglicano (apenas 10 por cento da parede celular total), circundada por um envelope externo contendo lipopolissacarídeos (LPS) e lipoproteínas. Este envelope externo é algumas vezes referido como uma segunda bicamada lipídica. A química desse envelope externo é muito diferente, entretanto, daquela da bicamada lipídica típica que forma as membranas plasmáticas das células.

Parede celular das bactérias
Parede celular das Archaea
As paredes celulares arqueais não apresentam peptidioglicano. Existem quatro tipos diferentes de paredes celulares arqueais. Um tipo é composto de pseudopeptidioglicano, que é semelhante ao peptidioglicano na morfologia, mas contém açúcares diferentes na cadeia polissacarídica.
Os outros três tipos de paredes celulares são compostos de polissacarídeos, glicoproteínas ou proteína pura.
Trabalhos recentes publicados a partir de 2014, mostram que existe no mínimo 5 tipos de paredes celulares presentes nas aqueias:
Tipo 1
O tipo mais comum de parede celular arqueal é uma camada S composta por
proteína ou glicoproteína. Com espessura: 20-40nm de espessura.
As paredes celulares com camada S estão presentes em alguns metanógenos como Methanococcus, Halophiles como Halobacterium e termófilos extremos como Sulpholobus, Pyrodictium etc.
Tipo 2
Neste tipo, camadas adicionais de material estão presentes fora da camada S. Em Methanosprillum, há uma bainha de proteína externa à camada S.
Tipo 3
Em Methanosarcina, a camada S é coberta por um material semelhante à condroitina chamado de mathanocondroitina.
Tipo 4
Em alguns Archaea como Methanothermus e Methanopyrus, a camada S é a camada mais externa e é separada da membrana plasmática por uma camada de moléculas semelhante a Peptidoglicano chamada psuedomureina.
Psuedomureina difere do Peptidoglicano por ter ácido N-acetiltalosaminurônico em vez de ácido N-acetil murâmico, L-aminoácidos em vez de D-aminoácidos que fazem ligações cruzadas e ligações glicosídica Beta (1->3) em vez de ligação glicosídica Beta (1->4) (Cortelazzo s/d).
Tipo 5
Neste tipo, a camada S está ausente. Em vez disso, a parede celular arqueal é de espessura camada única, espessa e homogênea semelhante a bactérias Gram-positivas. Essas archaea geralmente coram-se gram-positivas. Este tipo está presente em Methanobacterium, Halococcus etc.

Parede celular das arquéias
Reprodução assexuada
Os meios de reprodução assexuada usados por Archaea incluem reprodução binária, fissão múltipla, fragmentação ou brotamento.
O processo de divisão celular é controlado pelo ciclo celular; os cromossomos dentro das Archaea são replicados para produzir dois cromossomos filhos.
Archaea normalmente tem um único cromossomo circular. Os dois cromossomos filhos são então separados e a célula se divide. Este processo em Archaea parece ser semelhante aos sistemas bacterianos e eucarióticos.
Os cromossomos circulares contêm múltiplas origens de replicação, usando DNA polimerases que se assemelham a enzimas eucarióticas. No entanto, as proteínas envolvidas que direcionam a divisão celular são semelhantes às dos sistemas bacterianos.
A replicação do DNA, semelhante em todos os sistemas, envolve iniciação, alongamento e terminação. A replicação do DNA, começando nas origens de replicação presentes nos cromossomos circulares, requer proteínas iniciadoras. O recrutamento de proteínas adicionais por meio das proteínas iniciadoras permite a separação do DNA circular e resulta na formação de uma bolha.
O sistema de replicação de DNA em Archaea, semelhante a todos os sistemas, requer um grupo 3'OH livre antes que a síntese seja iniciada.
A primase usada para sintetizar um primer curto de RNA do grupo 3'OH livre varia dentro dos Archaea quando comparada com a de sistemas bacterianos e eucarióticos.
A primase usada por archaea é uma versão altamente derivada do motivo de reconhecimento de RNA (RRM). É estruturalmente semelhante às polimerases de RNA dependentes de RNA viral, transcriptases reversas, ciclases geradoras de nucleotídeos cíclicos e polimerases de DNA envolvidas na replicação e reparo do DNA. Uma vez que a RNA primase tenha realizado seu trabalho, a síntese de DNA continua de maneira semelhante pela qual o sistema eucariótico e o DNA são replicados
Adaptações das arquéas
As arquéas são principalmente anaeróbicos e vivem em ambientes com baixo teor de oxigênio. A maioria das arquéas não pode ser cultivada em laboratórios e, portanto, deve ser identificada por meio de técnicas independentes de cultura.
Os organismos neste domínio podem compartilhar algumas características com bactérias e eucariotos. Eles não apresentam núcleo, como as bactérias, mas compartilham alguns genes, vias metabólicas e enzimas com os eucariotos.
No entanto, esses organismos também têm algumas características únicas. Os lipídios da membrana das arquéas contêm ácido graxo ligado à molécula de glicerol por ligação de éter, em vez de ligação de éster, como nas bactérias e eucariotos.
Como as archaea habitam ambientes extremos diferentes, elas tendem a ter vias metabólicas distintas, bem como genes que sustentam sua sobrevivência.
As arquéas halofílicas (que toleram altas concentrações de sais) possuem um conjunto único de genes que limitam a extensão da osmose, facilitando sua sobrevivência.
A reprodução em archaea é assexuada por brotamento, fissão e fragmentação. O processo usual de divisão da mitose e meiose não ocorre.
A maioria das arquéas auxilia no processo de ciclagem biogeoquímica de vários elementos como carbono, nitrogênio e enxofre, etc.

Nutrição das arquéas (WP)
As arquéas são minúsculas, geralmente com menos de um mícron de comprimento (um milésimo de milímetro). Mesmo sob um microscópio óptico de alta potência, as maiores arqéas parecem minúsculos pontos. Felizmente, o microscópio eletrônico pode ampliar até mesmo esses minúsculos seres o suficiente para distinguir suas características físicas. Você pode ver as imagens, feitas usando uma variedade de técnicas micrográficas.
Você pode pensar que organismos tão pequenos não teriam muita variedade de formato ou forma, mas na verdade os formatos das arquéas são bastante diversos. Alguns são esféricos, uma forma conhecida como cocos, e podem ser perfeitamente redondos ou lobulados e protuberantes. Alguns são em forma de bastão, uma forma conhecida como bacilo, e variam de bastonetes curtos em forma de barra a formas longas e delgadas em forma de cabelo. Algumas espécies estranhas foram descobertas com uma forma triangular, ou mesmo uma forma quadrada como um selo postal!

Formas básicas das arquéas
1) Methanococcus janaschii, um coccus uma forma com numerosos flagella fixados em um lado. 2) Methanosarcina barkeri, um cocus lobado sem flagelos.
3) Methanothermus fervidus, uma forma de bacilo curto sem flagella.
4) Methanobacterium thermoautotrophicum, uma forma de bacilo alongado.
(Fonte: ucmp)
A diversidade estrutural entre os arqueáis não se limita à forma geral da célula. Archaea pode ter um ou mais flagelos anexados a eles, ou pode não ter flagelos. Os flagelos são apêndices semelhantes a cabelos usados para se mover e são fixados diretamente na membrana externa da célula. Quando vários flagelos estão presentes, eles geralmente são anexados todos em um lado da célula. Outros apêndices incluem redes de proteínas às quais as células podem se ancorar em grandes grupos.

Comparação de um flagelo com os "flagelos" das arquéas.
Como as bactérias, as arquéas não têm organelas nem membranas internas e seu DNA circular existe como uma única molécula semelhante a um plasmídeo. No entanto, seus tRNAs têm várias características que diferem de todos os outros seres vivos. As moléculas de tRNA ("RNA de transferência ou transportador") são importantes na decodificação da mensagem do DNA e na construção de proteínas.
Certas características da estrutura do tRNA são as mesmas em bactérias, plantas, animais, fungos e todos os seres vivos conhecidos, exceto Archaea.
Existem características no tRNA de arquéas que se parecem mais com organismos eucarióticos do que com bactérias, o que significa que as arquéas compartilham certas características em comum com você e não com as bactérias.
O mesmo é verdade para seus ribossomos, as moléculas gigantes de processamento que montam as proteínas para a célula. Embora os ribossomos bacterianos sejam sensíveis a certos agentes inibidores químicos, os ribossomos das arquéias e eucarióticos não são sensíveis a esses agentes. Isso pode sugerir uma relação próxima entre Archaea e eucariotos.
Tal como acontece com outras organismos vivos, as células das Archaeas têm uma membrana celular externa que serve como uma barreira entre a célula e seu ambiente. Dentro da membrana está o citoplasma, onde ocorrem todas funções vitais da arquéia e onde o DNA está localizado.
Em torno do lado externo de quase todas as células de arquéa há uma parede celular, uma camada semirrígida que ajuda a célula a manter sua forma e equilíbrio químico. Todas essas três regiões podem ser distinguidas nas células das eubactérias e na maioria dos outros seres vivos, mas quando você olha mais de perto cada região, descobre que as semelhanças são meramente estruturais, não químicas.
Em outras palavras, as Archaea constroem as mesmas estruturas que outros organismos, mas a partir de componentes químicos diferentes. Por exemplo, as paredes celulares de todas as eubactérias contêm o composto químico peptidoglicano.
As paredes celulares das arquéas não contêm esse composto, embora algumas espécies contenham um semelhante. Da mesma forma, as arquéas não produzem paredes de celulose (como as plantas) ou quitina (como os fungos). A parede celular dos arqueanos é quimicamente distinta.

Estrutura básica das arquéas: as três regiões básicas de uma célula de arquéa são o citoplasma, a membrana plasmática e a parede celular. Na figura, essas três regiões são rotuladas, com um aumento à direita da estrutura da membrana celular. As membranas das células das arquéas são quimicamente diferentes de todos os outros seres vivos, incluindo uma molécula de glicerol "reversa" e derivados de isopreno no lugar de ácidos graxos.(Fonte: ucmp)
As diferenças químicas mais marcantes entre Archaea e outros seres vivos estão em sua membrana celular. Existem quatro diferenças fundamentais entre a membrana arqueana e as de todas as outras células:
(1) quiralidade do glicerol,
(2) ligação de éter,
(3) cadeias isoprenóides e
(4) ramificação de cadeias laterais.
Podem parecer diferenças complexas, mas uma pequena explicação tornará as diferenças compreensíveis. (ucmp)
(1) Quiralidade do glicerol
A unidade básica a partir da qual as membranas celulares são construídas é o fosfolipídeo. Esta é uma molécula de glicerol que tem um fosfato adicionado em uma extremidade e duas cadeias laterais conectadas na outra extremidade. Quando a membrana celular é montada, a extremidade de glicerol e fosfato das moléculas ficam penduradas na superfície da membrana, com as longas cadeias laterais ensanduichadas no meio (veja a ilustração acima). Essa estratificação cria uma barreira química eficaz ao redor da célula e ajuda a manter o equilíbrio químico.
O glicerol usado para fazer fosfolipídios das arquéas é um estereoisômero do glicerol usado para construir membranas bacterianas e eucarióticas. Duas moléculas que são estereoisômeros são imagens espelhadas uma da outra. Coloque as mãos à sua frente, com as palmas para cima. Ambas as mãos estão orientadas com os dedos apontando para longe de você, os pulsos em sua direção e com as palmas para cima. No entanto, seus polegares apontam para direções diferentes porque cada mão é uma imagem espelhada da outra. Se você virar uma das mãos de modo que os dois polegares apontem da mesma forma, aquele não ficará mais com a palma para cima. (ucmp)
Esta é a mesma situação dos estereoisômeros do glicerol. Existem duas formas possíveis da molécula que são imagens espelhadas uma da outra. Não é possível transformar um no outro simplesmente girando-o. Enquanto as bactérias e os eucariotos têm D-glicerol em suas membranas, os Archaea têm L-glicerol nas suas. Isso é mais do que uma diferença geométrica. Os componentes químicos da célula precisam ser construídos por enzimas, e a "habilidade manual" (quiralidade) da molécula é determinada pela forma dessas enzimas. Uma célula que constrói uma forma não será capaz de construir a outra forma.(ucmp)


(2) Ligação de éter
Quando as cadeias laterais são adicionadas ao glicerol, a maioria dos organismos as unem usando uma ligação de éster (veja o diagrama acima). A cadeia lateral adicionada tem dois átomos de oxigênio ligados a uma extremidade. Um desses átomos de oxigênio é usado para formar a ligação com o glicerol e o outro se projeta para o lado quando a ligação é feita. Em contraste, as cadeias laterais das Archaea são ligadas por meio de uma ligação de éter, que não possui aquele átomo de oxigênio saliente adicional. Isso dá ao fosfolipídio resultante propriedades químicas diferentes dos lipídios da membrana de outros organismos. (ucmp)
(3) Cadeias de isoprenóides
As cadeias laterais nos fosfolipídios de bactérias e eucariotos são ácidos graxos, cadeias de geralmente 16 a 18 átomos de carbono. Os Archaea não usam ácidos graxos para construir seus fosfolipídios de membrana. Em vez disso, eles têm cadeias laterais de 20 átomos de carbono construídas a partir de isopreno.
O isopreno é o membro mais simples de uma classe de produtos químicos chamados terpenos. Por definição, um terpeno é qualquer molécula construída conectando moléculas de isopreno entre si, como se construísse com blocos de Lego®. Cada unidade de isopreno tem uma "cabeça" e uma "cauda" (novamente como um bloco de Lego®), mas ao contrário de suas contrapartes de brinquedo, os blocos de isopreno podem ser unidos de várias maneiras. Uma cabeça pode ser presa a uma cauda ou a outra extremidade da cabeça, e as caudas podem ser unidas da mesma forma. A imensa variedade de compostos de terpeno que podem ser construídos a partir de unidades simples de isopreno incluem beta-caroteno (uma vitamina), borrachas naturais e sintéticas, óleos essenciais de plantas (como hortelã) e hormônios esteróides (como estrogênio e testosterona).
Archaeol é um dos principais lipídios da membrana central das arquéias. Uma das principais características que distinguem as archaea das bactérias e dos eucariotos, como foi dito, são seus lipídios de membrana, onde o arqueol desempenha um papel importante. Por causa disso, o archaeol também é amplamente usado como um biomarcador para a atividade de arquéias antigas, especialmente os metanógenos.
Archaeol é geralmente composto pela ligação de duas cadeias de phytanyl ou fitanila às posições sn-2 e sn-3 de uma molécula de glicerol. As cadeias laterais altamente ramificadas são especuladas para explicar a permeabilidade muito baixa da membrana à base de arqueol, que pode ser uma das principais adaptações das arquéias a ambientes extremos.
(4) Ramificação das cadeias laterais
Não apenas as cadeias laterais das membranas arqueáis são construídas a partir de componentes diferentes, mas as próprias cadeias têm uma estrutura física diferente. Como o isopreno é usado para construir as cadeias laterais, existem ramificações laterais da cadeia principal (veja o diagrama acima). Os ácidos graxos de bactérias e eucariotos não possuem esses ramos laterais (o melhor que eles conseguem fazer é uma ligeira curvatura no meio), e isso cria algumas propriedades interessantes nas membranas arqueadas.
Por exemplo, as cadeias laterais de isopreno podem ser unidas. Isso pode significar que as duas cadeias laterais de um único fosfolipídeo podem se unir, ou podem ser unidas às cadeias laterais de outro fosfolipídeo no outro lado da membrana. Nenhum outro grupo de organismos pode formar tais fosfolipídios transmembrana.
Outra propriedade interessante dos ramos laterais é a capacidade de formar anéis de carbono. Isso acontece quando uma das ramificações laterais se enrola e se liga a outro átomo na cadeia para formar um anel de cinco átomos de carbono. Acredita-se que tais anéis forneçam estabilidade estrutural à membrana, uma vez que parecem ser mais comuns entre as espécies que vivem em altas temperaturas. Eles podem funcionar da mesma forma que o colesterol nas células eucarióticas para estabilizar as membranas. É interessante notar que o colesterol é outro terpeno!(ucmp).




Posição dos organismos do reino Archaea

Árvore filogenética universal baseada nas comparações entre sequências de rRNA de diversos organismos, (adaptada de Woese et al., 1990), na qual é possível observar os três domínios da vida e suas respectivas posições filogenéticas. Os números nos ramos correspondem aos grupos dos domínios Bacteria (1. Thermotogales, 2. Flavobactérias e organismos relacionados, 3. Cianobactérias, 4. Bactérias púrpuras, 5. Gram-positivas e 6. Bactérias verdes e não sulfurosas) Archaea (7. Pyrodictium, 8. Thermoproteus, 9. Thermococcales, 10. Methanococcales, 11. Methanobacteriales, 12. Methanomicrobiales e 13. Halófilos extremos) e Eukarya: 14. Metazoa (animais), 15. Ciliados, 16. Plantas verdes, 17. Fungos, 18. Flagelados, 19. Microsporídeos) (modif. de Ambrósio, 2016).
Esses organismos geralmente habitam ambientes extremos, como aberturas de fumarolas no fundo do mar, águas salinas, fontes termais e até mesmo abaixo de depósitos de petróleo.
O primeiro grupo de arqueias estudado foi o das "bactérias metanogênicas ou metanógenas".
A metanogênese foi descoberta no lago Maior na Itália em 1776, ao observar nele o burbulhar do "ar combustível".
Em 1882 observou-se que a produção de metano no intestino dos animais devia-se à presença de micro-organismos (Popoff, Tappeiner e Hoppe-Seyler).
Em 1936, ano que marcou o princípio da era moderna no estudo da metanogênese, H.A Barker oferece as bases científicas para o estudo da sua fisiologia e conseguiu desenvolver um meio de cultivo apropriado para o crescimento dos metanógenos. Nesse ano foram identificados os géneros Methanococcus e Methanosarcina.
As primeiras arqueias extremófilas foram encontradas em ambientes quentes. Em 1970, Thomas D. Brock, da Universidade de Wisconsin, descobriu a Thermoplasma, uma arqueia termoacidófila, e em 1972 a Sulfolobus, uma hipertermófila.
Brock se iniciou em 1969 no campo da biologia dos hipertermófilos com a descoberta de Thermus aquaticus, que não é uma arquea mas uma eubactéria.
Em 1977 identificam-se as arqueias como o grupo procarionte mais distante das eubactérias ao descobrir que os metanógenos apresentam uma profunda divergência com todas as bactérias estudadas. Nesse mesmo ano foi proposta a categoria de super-reino para este grupo com o nome de Archaebacteria. Atualmente as Archaea estão no seu próprio reino, o reino Archaea, dentro de um domínio próprio o Domínio Archaea.

Os três domínios dos organismos vivos. Bactérias e Archaea são procariotos, mas diferem o suficiente para serem colocadas em domínios separados e distintos. Acredita-se que um ancestral das modernas Archaea deu origem ao Eukarya, o terceiro domínio da vida. Alguns grupos de Archaea e Bacteria são mostrados.
As arquéas, que há algum tempo atrás eram chamadas de arqueobactérias, e estavam colocadas juntos com as Eubactérias (num mesmo reino, Monera), são organismos procariontes pertencentes ao domínio próprio o domínio Archaea.
Elas apresentam um conjunto de características que justifica serem incluídas em um reino distinto do reino das Eubactérias.
Maiores detalhes AQUI.

Micrografia eletrônica de transmissão colorida artificialmente (TEM) mostra uma seção através da Archaebacterium Methanococcoides burtonii. Esta arquéia psicrofílica (que gosta de frio) foi descoberta em 1992 em Ace Lake, na Antártida, e pode sobreviver em temperaturas tão baixas quanto -2,5 graus Celsius. Como uma bactéria metanogênica, é capaz de formar metano a partir de dióxido de carbono e hidrogênio.
(by DR M.ROHDE, GBF/Science Photo Library/Getty Images).
[This is a colored transmission electron micrograph (TEM) of a section through the Archaebacterium Methanococcoides burtonii. This psychrophilic (cold-loving) Archaebacterium was discovered in 1992 in Ace Lake, Antarctica, and can survive in temperatures as low as -2.5 degrees Celsius. As a methanogenic bacterium, it is able to form methane from carbon dioxide and hydrogen. DR M.ROHDE, GBF/Science Photo Library/Getty Images].
Methanococcoides burtonii Franzmann et al., 1993, é um archaea extremófila da família Methanosarcinaceae, uma família de três gêneros de arquéias com células em forma de cocos.
Methanococcoides burtonii adaptou-se à vida na Antártida, onde reside no Lago Ace em temperaturas que permanecem permanentemente entre 1 e 2 °C.
M. burtonii foi descoberto pela primeira vez por um limnologista austríaco chamado Harry Burton. Foi determinado que a temperatura ideal de crescimento era em torno de 23°C.
M. burtonii é capaz de crescer em substratos metilados e tolera uma ampla faixa de temperaturas de crescimento (< 4° a 29°C). A adaptação ao frio em M. burtonii envolve mudanças específicas na insaturação lipídica da membrana e proteínas flexíveis. M. burtonii são cocos irregulares, com tamanho aproxcimado de 1,8 micrômetros de diâmetro. Essas arquéias ocorrem isoladamente ou em pares. Quando se utiliza a coloração de Gram, as células sofrem lise; eles também se lisam em soluções hipotônicas. M. burtonii são móveis com um único flagelo e não possuem estruturas de armazenamento nem organelas com membranas internas no citoplasma como todo procarioto. Além disso M. burtonii são archaeas formadoras de colônias, geralmente ocorrendo em colônias <1 milímetro que são circulares e convexas.
As células de M. burtonii fluorescem em azul quando expostas à iluminação UV. O pH inicial ideal para crescimento é 7,7.

Methanococcoides brutonii
As células de M. burtonii mostraram-se resistentes à penicilina, ampicilina, tetraciclina, vancomicina e eritromicina. Embora tenha evoluído a capacidade de se sustentar em ambientes considerados "extremofílicos" para archaea (1-2 °C), M. burtonii cresce de forma ideal a 23 °C. Essa espécie é um metanógeno obrigatoriamente metilotrófico capaz de usar metilaminas e metanol, mas não formiato, H2CO2 ou acetato para seu crescimento. O metano é um gás de efeito estufa, e os metanogênicos desempenham um papel crítico no aquecimento global e no ciclo global do carbono por meio da produção de metano.
Os mecanismos pelos quais as células se adaptam ao frio foram obtidos principalmente a partir da resposta ao choque frio em procariontes mesófilos no domínio filogenético Bactérias.
A seqüência do genoma do metanogênico psicrotolerante (tolerante ao frio) M. burtonii, pelo "DOE Joint Genome Project", permitirá aos pesquisadores identificar os genes necessários à sobrevivência e adaptação dessas Archaea ao frio. Até o momento, nenhum genoma foi concluído para quaisquer organismos adaptados ao frio e o sequenciamento do genoma de M. burtonii fornecerá a primeira visão abrangente sobre a composição genética necessária para a adaptação ao frio.
Pela análise de sequências comparativas com genomas metanogênicos, incluindo o mesófilo Methanosarcina barkeri (Tmax: 40°C), o termófilo Methanosarcina thermophila (Tmax: 55°C) e o termófilo extremo Methanococcus jannaschii (Tmax: 85°C) será possível identificar um subconjunto de genes exclusivos deste metanógeno adaptado ao frio. As comparações de estrutura de proteínas in silico também podem destacar estratégias adaptativas.
Além das implicações evolutivas inerentes à comparação de mecanismos de adaptação ao frio de archaea com aqueles em Bacteria e Eukarya, a conclusão da sequência do genoma de M. burtonii expandirá nossa compreensão da biocomplexidade orgânica e metabólica que evoluiu em associação com microrganismos adaptados ao frio. Completar a sequência genômica desse extremófilo contribuirá para o objetivo de longo prazo de estabelecer as estratégias globais de adaptação ao frio dos microrganismos e definir os limites da vida na Terra e em ambientes extraterrestres. (genome)

Micrografia eletrônica de varredura de L. aggregans. Fonte: Johannes Sikorski/Researchgate.
FORMA
Basic Archaeal Shapes
1) Methanococcus janaschii, a coccus form with numerous flagella attached to one side. 2) Methanosarcina barkeri, a lobed coccus form lacking flagella.
3) Methanothermus fervidus, a short bacillus form without flagella.
4) Methanobacterium thermoautotrophicum, an elongate bacillus form.
Formas archaeáis básicas: 1) Methanococcus janaschii, uma forma coccus com numerosos flagelos anexados a um lado. 2) Methanosarcina barkeri, uma forma de cocos lobados sem flagelos. 3) Methanothermus fervidus, um bacilo curto sem flagelo. 4) Methanobacterium thermoautotrophicum, uma forma alongada de bacilo
Vivendo no extremo
Em geral, o domínio das Archaea é dividido em 3 principais grupos de acordo com as características dos seus integrantes:
Hipertermófilos
Habitam ambientes com altas temperaturas ou temperaturas muito baixas. Algumas arqueas são termoacidófilas, tolerando ambientes que possuem tanto alta temperatura como extrema acidez. Exemplo: hipertermófilo Ignisphaera aggregans.
Metanógenos
Organismos que produzem metano como subproduto do metabolismo energético. Vivem apenas em ambientes livres de oxigênio (anaeróbios). Exemplo: Methanobrevibacter smithii.
Halófilos extremos
Procariotos que habitam ambientes hipersalinos. Podem ser encontrados em lagos com altíssima concentração de sal, como o Mar Morto. Exemplo: Halobacterium salinarum.
Micrografia eletrônica de varredura de Ignisphaera aggregans.
(Fonte: Johannes Sikorski/Researchgate)
Extremófilos
Muitas espécies de archaea são metanogênicas que utilizam respiração celular anaeróbica para produzir metano como subproduto.
Mesmo que a fotossíntese geradora de oxigênio não ocorra nesses organismos, alguns deles (fototróficos) usam a luz solar como fonte de energia.


Archaea foram encontrados em fontes termais vulcânicas. A foto mostra a Grande Fonte Prismática do Parque Nacional de Yellowstone EUA.
A maioria das arqueas são autotróficas quimiossintetizantes, enquanto as bactérias podem apresentar diversificados modos de vida.
Essa característica permite que elas vivam em locais extremos, onde muitos outros organismos não sobreviveriam. Geralmente esses organismos apresentam características que não só os permitem sobreviver nesses ambientes, como também crescem e se multiplicam melhorar em ambientes extremos.
Devido a isso, eles são chamados de extremófilos (do latim: extremus, extremo e φιλία, philiā, que significa amante, i.e., são amantes de condições extremas. Estes organimos podem ser divididos em:
Halófilos extremos
São organismos que vivem em ambientes extremamente salinos, como o Mar Morto, em Israel. Um exemplo são indivíduos do gênero Halobacterium sp.
Termófilos extremos
Os termófilos extremos são organismos que vivem em ambientes extremamente quentes. Esses organismos vivem próximo de fontes termais submarinas e utilizam a energia química do gás sulfídrico para sintetizar compostos orgânicos. Um exemplo são os chamados de “cepa 121”, pois se reproduzem nesses ambientes a uma temperatura de 121°C. Eles possuem adaptações que tornam suas proteínas e material genético estáveis nessas temperaturas.
Embora sejam conhecidos por viverem em regiões extremas, alguns organismos vivem em ambientes moderados, como pântanos e vísceras de herbívoros, sendo chamados de metanogênicos, pois utilizam hidrogênio para reduzir o gás carbônico e produzir metano.
Divisões do reino Archaea

Divisão do reino Archaea
Euryarchaeota
Este filo inclui metanógenos, que produzem metano como um produto residual metabólico, e halobactérias, que vivem em um ambiente extremamente salino.
Metanógenos
A produção de metano causa flatulência em humanos e outros animais.
Halobactérias: grandes flores desta arquéia amante do sal aparecem avermelhadas devido à presença de bacteriorodopsina na membrana. A bacteriorodopsina está relacionada ao pigmento da retina rodopsina.

Crenarchaeota
Os membros desse filo onipresente desempenham um papel importante na fixação de carbono. Muitos membros desse grupo são extremófilos dependentes de enxofre. Alguns são termofílicos ou hipertermofílicos.

Imagem de Microscopia Eletrônica de Varredura de Venenivibrio stagnispumantis, uma espécie que ganha energia oxidando o gás hidrogênio. (courses)
Os Crenarchaeota que foram cultivados e caracterizados são termofílicos. Muitos também são acidofílicos. Eles ocorrem em fontes termais ácidas e lamaçais e em fontes hidrotermais submarinas (thermal vents) ou fumarolas. As temperaturas de crescimento podem exceder 100 °C onde a pressão hidrostática impede a ebulição; por exemplo, para a espécie Pyrolobus fumarii, a temperatura ótima é de 106 °C. A maioria dos Crenarchaeota são anaeróbicas.
Para o metabolismo gerador de energia, a respiração que faz uso do enxofre elementar (S°) é comum, embora muitos outros aceptores de elétrons, incluindo o oxigênio (O2), sejam usados por várias espécies. O gás hidrogênio (H2) de origem geotérmica costuma servir como doador de elétrons. Muitas espécies também podem usar compostos orgânicos e, para aeróbios, S° também pode ser um doador de elétrons. Muitos Crenarchaeota têm formas celulares incomuns, incluindo filamentos e discos irregulares, embora bastões e cocos irregulares sejam comuns. Três ordens são reconhecidas: os Thermoproteales, os Desulfurococcales e os Sulfolobales.

Thermoproteus uzoniensis sp. nov., uma nova arqueobactéria extremamente termofílica das fontes termais continentais de Kamchatka.
Muitos Crenarchaeota hipertermofílicos têm matrizes cristalinas bidimensionais de subunidades (glico-)proteínas (a camada S) como o componente mais rígido de suas paredes celulares. Na maioria dos casos, essas matrizes de proteínas constituem as superfícies mais externas das células. As próprias subunidades são diretamente ancoradas na membrana plasmática por estruturas semelhantes a talos. O espaço entre a MP e a camada S é chamado de "espaço quase-periplasmático" por analogia às estruturas equivalentes nos parede celular (envelopes) de células bacterianas típicas.
Ignicoccus é um archaeon hipertermofílico pertencente à subdivisão Desulfurococcales de Crenarchaeota. É um organismo quimiolitoautotrófico que obtém sua energia pela redução do enxofre elementar com o hidrogênio molecular. As células de Ignicoccus foram examinadas ultraestruturalmente após cultivo em capilares de celulose e processamento por congelamento de alta pressão. Eles consistentemente mostraram uma estrutura de envelope celular até então desconhecida entre as arquéias: MP e ME (membrana externa) separados por um espaço periplasmático de largura variável (20–200 nm) contendo vesículas ligadas à membrana. A bainha externa, de aproximadamente 10 nm de largura, parecia se assemelhar aos MEs de bactérias Gram-negativas.
A bainha de Ignicoccus contém três tipos de partículas: (1) numerosas partículas individuais empacotadas irregularmente, com cerca de 8 nm de diâmetro; (2) poros putativos com um diâmetro de 24 nm; e (3) partículas minúsculas dispostas em um anel com um diâmetro de 130 nm circundando os poros. Aglomerados de até oito partículas, cada partícula de 12 nm de diâmetro, foram conspícuos. As células congeladas exibiam uma superfície lisa sem um padrão regular, com planos de fratura freqüentes na bainha externa. Esta observação indicou aos pesquisadores a presença de um ME e a ausência de uma camada S. Estudos tem ilustrado a nova arquitetura complexa do envelope celular de Ignicoccus.
(Também foram descobertos virus que infectam as espécie de Crenarchaeota) (SAIER JR, 2009).
Sulfolobus sp
Os membros deste gênero crescem em fontes vulcânicas em temperaturas entre 75º e 80º C e em pH entre 2 e 3.
Nanoarchaeota
Este grupo atualmente contém apenas uma espécie: Nanoarchaeum equitans. A espécie Nanoarchaeum equitans foi coletada e isolada do fundo do Oceano Atlântico e de uma fonte hidrotermal no Parque Nacional de Yellowstone. É um simbionte obrigatório com Ignicoccus, outra espécie de arquéia.
Korarchaeota
Este grupo é considerado uma das formas de vida mais primitivas. Membros deste filo só foram encontrados no Obsidian Pool, uma fonte termal no Parque Nacional de Yellowstone. Nenhum membro desta espécie foi cultivado.
No domínio Archaea, existem três divisões ou filos principais. São eles: Crenarchaeota,
Euryarchaeota e
Korarchaeota.
Crenarchaeota
Os Crenarchaeota consistem principalmente de hipertermófilos e termoacidófilos. Os microrganismos hipertermofílicos vivem em ambientes extremamente quentes ou frios.
Os termoacidófilos são organismos microscópicos que vivem em ambientes extremamente quentes e ácidos. Seus habitats têm um pH entre 5 e 1. Você encontrará esses organismos em fontes hidrotermais e fontes termais.
Espécies de Crenarchaeota Exemplos de Crenarchaeotas incluem:
Sulfolobus acidocaldarius
Encontrado próximo a ambientes vulcânicos em fontes quentes e ácidas contendo enxofre.
Pyrolobus fumarii
Vive em temperaturas entre 90 e 113ºC.
Euryarchaeota
Os organismos Euryarchaeota consistem principalmente de halófilos extremos e metanógenos. Organismos halofílicos extremos vivem em habitats salgados. Eles precisam de ambientes salgados para sobreviver. Você encontrará esses organismos em lagos salgados ou áreas onde a água do mar evaporou.

Este extremófilo de gênero desconhecido (provavelmente, Halomicrobium sp. Ou Halobacterium sp.) Foi encontrado crescendo em alta salinidade (2,5 vezes as concentrações na água do mar normal). É um exemplo de variações dramáticas no tamanho ou forma da célula, denominado pleomorfismo. Halobacteriales são uma ordem de Halobacteria encontrada em água saturada ou quase saturada com sal e são comuns onde quer que sejam encontradas grandes quantidades de sal, umidade e material orgânico. Halobactérias requerem oxigênio e são incapazes de fixar carbono do dióxido de carbono, mas podem sobreviver em ambientes salgados por causa de sua maneira única de criar energia por meio da fotossíntese, que é diferente de outras formas de fotossíntese que usam clorofila. Grandes florações de halobactérias aparecem avermelhadas, devido ao pigmento bacteriorodopsina. Este pigmento é usado para absorver luz, que fornece energia para criar ATP. Um segundo pigmento, a halorodopsina, é usado para bombear íons de cloreto em resposta aos fótons, criando assim um gradiente de voltagem e auxiliando na produção de energia da luz. (treeoflife)
Os metanógenos requerem condições livres de oxigênio para sobreviver, pois são anaeróbicos. Eles produzem gás metano como um subproduto do metabolismo. Você encontrará esses organismos em ambientes como pântanos, locais muito úmidos (charcos), lagos de gelo, no intestino de animais (vacas, veados, humanos) e esgoto.

 1. PNAS, 87:4576-79, 1990; 2. Trends Microbiol, 19:580-87, 2011; 3. Nature, 417:63-67, 2002; 4. Nature, 499:431-37, 2013; 5. Nature, 521:173-79, 2015; 6. Nature, 541:353-58, 2017
1. PNAS, 87:4576-79, 1990; 2. Trends Microbiol, 19:580-87, 2011; 3. Nature, 417:63-67, 2002; 4. Nature, 499:431-37, 2013; 5. Nature, 521:173-79, 2015; 6. Nature, 541:353-58, 2017
Exemplos de Euryarchaeota
Halobacterium
Nesse táxon estão incluídos várias espécies de organismos halofílicos que são encontrados em lagos salgados e ambientes oceânicos de alta salinidade.
Gênero Methanococcus
Methanococcus jannaschii foi o primeiro arquéia sequenciado geneticamente. Este metanógenio vive perto de fontes hidrotermais.
Methanococcoides burtonii esse metanógenos psicrofílicos (ψυχρός: psychrós frio, gelo, e φίλος: phílos, amigo, amantes do frio) foram descobertos na Antártica e podem sobreviver a temperaturas extremamente frias.

Methanococcus jannaschii
Methanococcus jannaschii (também conhecido como Methanocaldococcus jannaschii) foi descoberto vivendo em águas extremamente quentes e privadas de oxigênio na base de uma chaminé de fumaça branca no fundo do oceano, e foi o primeiro Archaea a ter seu genoma totalmente sequenciado. A maioria de seus genes relacionados à produção de energia, divisão celular e metabolismo foram considerados mais semelhantes aos encontrados em bactérias, enquanto aqueles relacionados à transcrição, tradução e replicação revelaram-se mais semelhantes aos gnes encontrados em eucariotos.
Methanococcus é um gênero de metanógenos, i.e., produtores de metano. Os metanogênicos são extremamente importantes em ambientes anaeróbios (com falta de oxigênio) porque, à medida que convertem compostos orgânicos em metano, eles fornecem caminhos para que compostos existentes em ambientes anaeróbios escapem para a atmosfera. Methanococcus jannaschii produz muitos cofatores, coenzimas e enzimas exclusivos durante a metanogênese, macromoléculas que podem ser valiosas em aplicações de biotecnologia.
Os ambientes frios em que os psicrófilos habitam são onipresentes na Terra, pois uma grande fração de nossa superfície planetária experimenta temperaturas inferiores a 15°C. Eles estão presentes no permafrost, no gelo polar, nas geleiras, campos de neve e águas profundas do oceano. Esses organismos também podem ser encontrados em bolsões de gelo marinho com alto conteúdo de salinidade. A atividade bacteriana foi medida em solos congelados abaixo de -39 °C.
Korarchaeota
Os organismos Korarchaeota são considerados formas de vida muito primitivas. Atualmente, pouco se sabe sobre as principais características desses organismos. Sabemos que são termofílicos e foram encontrados em fontes termais e piscinas de obsidiana.
Relacionamento filogenético
Archaea são organismos interessantes porque possuem genes semelhantes a bactérias e eucariotos. Filogeneticamente falando, acredita-se que arqueas e bactérias tenham se desenvolvido separadamente de um ancestral comum. Acredita-se que os eucariotos tenham se ramificado dos Archaea milhões de anos depois do surgimento daqueles. Isso sugere que os arqueanos estão mais intimamente relacionados aos Eukarya do que às bactérias.

EXERCÍCIOS
1) (Uerj 2011) A influência de fatores ambientais, como a disponibilidade de alimentos, sobre o crescimento dos seres vivos pode ser avaliada experimentalmente. Considere, por exemplo, um inóculo da bactéria E. coli que foi introduzido em um meio nutritivo adequado. O tempo de geração, ou seja, o intervalo de tempo necessário para que uma célula se duplique, foi medido durante a fase de crescimento exponencial e durante a fase estacionária. Observe os gráficos abaixo:
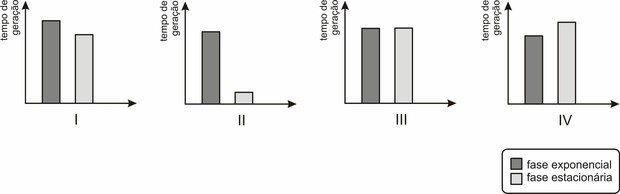
O resultado desse experimento, em relação à influência de fatores ambientais no crescimento bacteriano, está representado pelo gráfico de número:
(a) I
(b) II
(c) III
(d) IV
2) (Enem 2003) Na embalagem de um antibiótico, encontra-se uma bula que, entre outras informações, explica a ação do remédio do seguinte modo: O medicamento atua por inibição da síntese proteica bacteriana. Essa afirmação permite concluir que o antibiótico:
(a) impede a fotossíntese realizada pelas bactérias causadoras da doença e, assim, elas não se alimentam e morrem.
(b) altera as informações genéticas das bactérias causadoras da doença, o que impede manutenção e reprodução desses organismos.
(c) dissolve as membranas das bactérias responsáveis pela doença, o que dificulta o transporte de nutrientes e provoca a morte delas.
(d) elimina os vírus causadores da doença, pois não conseguem obter as proteínas que seriam produzidas pelas bactérias que parasitam.
(e) interrompe a produção de proteína das bactérias causadoras da doença, o que impede sua multiplicação pelo bloqueio de funções vitais.
Resolução
1) Resolução: na fase exponencial há disponibilidade de alimentos, com isso o tempo de geração de bactérias é menor, crescendo rapidamente a colônia bacteriana. Entretanto, na fase estacionária, há uma escassez de alimento, fazendo com que o tempo de geração seja menor, diminuindo o crescimento bacteriano. Letra D.
2) Resolução: a inibição da síntese proteica das baterias realizada pelos antibióticos, causa a morte desse microorganismo. Letra E.
REFERÊNCIAS
https://www.centerlab.com/blog/Centernews_123/ (01/III/2020)
http://jaguar.fcav.unesp.br/download/deptos/biologia/Manoel_Victor/genetica_de_bacterias.pdf (01/III/2020)
http://kentsimmons.uwinnipeg.ca/16cm05/1116/16monera.htm
http://www.enq.ufsc.br/labs/probio/disc_eng_bioq/trabalhos_pos2004/microorganismos/BACTERIAS.htm
http://www.enq.ufsc.br/labs/probio/disc_eng_bioq/trabalhos_pos2004/microorganismos/BACTERIAS.htm
http://amigonerd.net/trabalho/27634-pseudomona
http://www.enq.ufsc.br/labs/probio/disc_eng_bioq/trabalhos_pos2004/microorganismos/BACTERIAS.htm
http://amigonerd.net/trabalho/27634-pseudomona
https://www.sbemsp.org.br/imprensa/releases/119-endocrinologistas-alertam-para-uso-inadequado-do-lugol (visitado em março de 2020)
https://brasil.elpais.com/brasil/2020/01/17/ciencia/1579284583_584643.html
https://brasil.elpais.com/brasil/2020/01/17/ciencia/1579284583_584643.html
https://www.nature.com/articles/ismej201166 (visitado em 31/mar/2021)
http://aadityayadav648.blogspot.com/2013/ (visitado em 31/mar/2021)
https://pt.wikipedia.org/wiki/Elementos_gen%C3%A9ticos_m%C3%B3veis
(visitado em 09/março/2021)
(visitado em 09/março/2021)
(visitado em 09/março/2021)
(visitado em 09/março/2021)
Biorremediação com bactérias
WOESE, C.R., FOX G.E. (November 1977). Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proceedings of the National Academy of Sciences of the United States of America. 74 (11): 5088–90.
Raven, P.H., Evert, R.F., and Eichhorn, S.E. 1992. Biology of Plants. Fifth edition. Worth Publishers, New York, New York. pp. 602-611.
Clorofila e metabolismo vegetal
https://bio.libretexts.org/Bookshelves/Microbiology/Book%3A_Microbiology_(Boundless)/7%3A_Microbial_Genetics/7.08%3A_Archaeal_Genetics/7.8A%3A_Chromosomes_and_DNA_Replication_in_the_Archaea#:~:text=Archaea%20typically%20have%20a%20single,both%20bacterial%20and%20eukaryotic%20systems.
https://www.fcav.unesp.br/Home/departamentos/producaovegetal/everloncidrigobelo3646/estrutura-e-funcao-celular-em-bacterias-e-archaea.pdf
https://edisciplinas.usp.br/pluginfile.php/4649040/mod_resource/content/1/CAP%2028%20PAREDES%20CELULARES.pdf
https://www.fcav.unesp.br/Home/departamentos/producaovegetal/everloncidrigobelo3646/estrutura-e-funcao-celular-em-bacterias-e-archaea.pdf
https://courses.lumenlearning.com/boundless-microbiology/chapter/crenarchaeota/
https://courses.lumenlearning.com/boundless-microbiology/chapter/crenarchaeota/
Penicilinas
https://www.sanarsaude.com/portal/concursos/artigos-noticias/penicilinas-artigo-farmacia-tudo-que-voce-precisa-saber
Como não proceder
https://blog.gsuplementos.com.br/solucao-de-lugol/
Posição cientifica dos pesquisadores
https://www.sbemsp.org.br/imprensa/releases/119-endocrinologistas-alertam-para-uso-inadequado-do-lugol
Como não proceder
https://blog.gsuplementos.com.br/solucao-de-lugol/
Posição cientifica dos pesquisadores
https://www.sbemsp.org.br/imprensa/releases/119-endocrinologistas-alertam-para-uso-inadequado-do-lugol
(Atualizado em abril 2022)
0 comentários:
Postar um comentário