DIVISÃO DO FILO MOLLUSCA

Sépia, Sepia officinnalis L. é uma cefalópode de dotado de grande inteligência e habilidades além de um fantástico mecanismo de camuflagem.



Os polvos são predardes de sangue frio e têm todo um conjunto de ferramentas para subjugar diferentes tipos de presas. Com as amêijoas, eles agarram a concha com seus tentáculos e, usando uma língua afiada e cheia de dentes a rádula, fazem um pequeno orifício na concha e injetam um veneno paralisante. O molusco drogado relaxa seus músculos que mantém a concha fechada e o polvo pode facilmente forçar a abertura. Com o ex-inquilino agora já devorado, sua presença é apenas uma memória evanescente, o polvo assume a casa do molusco e usa essa concha grossa para se proteger de seus próprios predadores, mantendo um olho aberto para o perigo.
Introdução
Os moluscos formam um grupo monofilético morfologicamente muito bem definido. Apesar de sua grande diversidade de formas, alguns caracteres podem ser reconhecidos em quase todas as espécies de moluscos, e podem ser interpretados como ancestrais do filo. Caracteres compartilhados por um ancestral e todos os seus descentes são chamados de caracteres sinapomórficos.
Um dos métodos que tem sido utilizado mais freqüentemente para se buscar as relações de parentesco (ou filogenéticas) é o método cladístico. Esse método utiliza caracteres sinapomórficos, ou sinapomorfias como indicadores de parentesco.
Caracteres sinapomórficos (ou sinapomorfias) são caracteres homólogos derivados, herdados de um ancestral comum recente por dois ou mais táxons e que permitem reuni-los em grupos monofiléticos, que na maioria dos casos mostra um padrão ramificado.
O resultado é resumido em um gráfico, denominado cladograma, que apresenta a distribuição de caracteres. O cladograma é uma hipótese evolutiva. O cladograma é transformado em uma árvore filogenética, quando interpretado do ponto de vista temporal e quando assume-se que cada ponto de ramificação (nó) representa um ancestral, real ou hipotético e que cada ramo representa uma linhagem evolutiva de um dado grupo.
O molusco ancestral mais antigo provavelmente tinha três caracteres:
Manto (ou pálio): grande área de epitélio dorsal com uma espessa cutícula com espículas calcárias que recobre a massa visceral.
Pé: expansão musculosa ventral achatada e ciliada utilizada para o deslocamento rastejante ou outras formas de locomoção
Rádula: faixa cuticular com dentes formada em uma bolsa no epitélio ventral do esófago e utilizada para raspagem durante a alimentação. É considerada uma estrutura exclusiva dos moluscos.(portal)
Síntese filogenética dos Mollusca
Nos últimos anos, os estudos fundamentados em marcadores moleculares largamente usados em filogenia (e.g. histonas do DNA nuclear, proteínas afins das histonas de DNA mitocondrial, sequências genéticas baseadas em cDNA, sequências genéticas de 18S rRNA e 26S rRNA, sequências genéticas mitocondriais 16S rRNA) trouxeram novas pistas sobre a evolução e relações filogenéticas entre os bivalves e os outros moluscos (e.g. Giribet & Wheeler, 2002; Giribet & Distel, 2003; Giribet et al., 2006; Bieler et al., 2014).
Desta forma, considera-se hoje que o Filo Mollusca está dividido em nove classes (e.g. Salvini-Plawen, 2003; Haszprunar et al., 2008; Kocot et al., 2011; Smith et al., 2011, 2013; Telford & Budd, 2011), designadas respetivamente por:
Aplacophora von Ihering, 1876
Polyplacophora Gray, 1821;
Monoplacophora Odhner, in Wenz, 1940;
Gastropoda Cuvier, 1795;
Cephalopoda Cuvier, 1795;
Bivalvia Linnaeus, 1758;
Scaphopoda Bronn, 1862;
†Rostroconchia Cox, 1960 (Pojeta, Runnegar, Morris & Newell, 1972 (cambriano ao perminao) classe extinta.
Com ligação às relações filogenéticas, no seio do Filo Mollusca, vários cenários têm sido traçados para clados que traduzam as conexões entre as primeiras oito classes referidas, todas elas com representantes atuais. As mais generalizadas na literatura agrupam-nas em dois clados: Aculifera Hatschek, in Blumrich, 1891) e Conchifera Gegenbaur, 1878 (e.g. Kocot et al., 2011; Smith et al., 2011, 2013).



Dendrogramas mostrando o relacionamento entre as classes de moluscos
Desta forma, a Classe Bivalvia insere-se no clado Conchifera, sendo que a sua posição, dentro deste clado principal, varia consoante as metodologias de investigação utilizadas. Alguns trabalhos de filogenia genómica suportam um clado que reúne bivalves e gastrópodes, como taxa irmãos, denominado de Pleistomollusca (Kocot et al., 2011). A proposta de Smith et al. (2011) alterada por corrigenda em Smith et al. (2013), por seu turno, invoca a maior afinidade entre os gastrópodes e os escafópodes, colocando-os como taxa irmãos dentro do clado Conchifera.
Evolução e radiação dos Bivalvia O conhecimento paleontológico sobre a história evolutiva dos bivalves remonta ao primeiro impulso da denominada explosão câmbrica. Assim, no Fortuniano, embora com uma distribuição restrita, surgem os primeiros registos de bivalves, e.g. Fordilla troyensis Barrande, 1881 e F. Sibirica Krasilova, 1977.
Os Fordilla fizeram parte da denominada fauna tommotianada plataforma siberiana, que precedeu a primeira ocorrência conhecida de trilobitas, coevas com o bivalve Pojetaia runnegari Jell, 1980, este já com uma distribuição mais alargada que F. troyensis (Parkhaev, 2008, 2017).
Os bivalves do cambriano utilizariam o pé para locomoção e alimentação (Morton, 1996). As brânquias seriam simples e pequenas, servindo apenas para respirar e não desempenhariam qualquer papel na alimentação (Fang, 2006).
São referidos como sendo infaunais (e.g. Pojeta & Runnegar, 1985), ou epifaunais que se deslocariam lentamente sobre o tapete microbiano dos fundos marinhos do Cambriano (e.g. Cope & Kříž, 2013).
Apesar da dificuldade de análise e interpretação de muitos dos somatofósseis do cambriano atribuídos a bivalves, alguns dos quais vieram a ser reinterpretados em estudos subsequentes (e.g. Pojeta, 2000) como braquiópodes, rostroconchias ou outras formas com afinidade dúbia, a verdade é que os gêneros Fordilla, Pojetaia, Tuarangia MacKinnon, 1982, Camya Hinz-Schallreuter, 1995 e Arhouriella Geyer & Streng, 1998, reforçaram o seu estatuto de bivalves primitivos (Pojeta, 2000; Elicki & Gürsu, 2009).
Mais tarde, o Período Ordovciano foi caracterizado pela prevalência de condições geodinâmicas e paleoambientais muito particulares, que terão proporcionado um enorme incremento na biodiversidade e biocomplexidade da vida marinha.
Este evento, habitualmente designado por GOBE (Great Ordovician Biodiversification Event), terá ocorrido essencialmente durante o Ordoviciano Inferior e Médio, num intervalo de tempo de apenas 25 Ma, havendo autores que elencam o GOBE à enorme diversificação que ocorreu no Cambriano (e.g. Servais et al., 2009).
As condições paleoambientais favoráveis a esta radiação, de enorme amplitude na vida marinha, terão tido origem num elevado nível eustático (elevado nível marinho), referido como sendo o mais alto de todo o Paleozoico (e.g. Hallam, 1992; Munnecke et al., 2010; Haq & Schutter, 2008).
Eustasia é um termo usado para designar o processo que se traduz no movimento eustático, ou seja, na variação global relativa do nível do mar. A eustasia traduz-se em nível planetário por uma alteração do nível das águas do mar em relação ao nível da terra firme, causada por variações do volume de água no oceano global ou por variação do volume global das bacias oceânicas.
Segundo Haq e Schutter (2008, pp. 66-67), o máximo, cerca de 225 m acima do nível atual, terá ocorrido no Katiano (circa 449 Ma). Assim, terão existido grandes áreas cobertas por mares epicontinentais, de fundos rasos, com as maiores extensões de plataformas tropicais de todo o Fanerozoico (Munnecke et al., 2010), a que corresponderiam áreas emersas restritas relacionadas com arquipélagos emergentes.
Segundo Cocks e Torsvik (2002), estas áreas continentais encontravam-se fortemente dispersas como resultado da fragmentação da Rodínia (palavra russa que significa: Terra mãe) e a atividade magmática e tectónica teria sido persistente, com movimentos rápidos das placas tectónicas e atividade vulcânica generalizada. Nestas condições terá ocorrido um importante percurso evolutivo de diversificação dos bivalves, a partir de um pequeno grupo de paleotaxodontes cambrianos (Cope, 1996; Cope & Babin, 1999). No Ordoviciano terão surgido bivalves epifaunais (e.g. pterinídeos e modiomorfídeos) (Pojeta, 1971), com Waller (1978) a sustentar evidências de bivalves nadadores (limoides).
Foi justamente nessa época há cerca de 750 milhões de anos, tempo de mudanças climáticas intensas, que Rodínia começou a se fragmentar e a "dança" dos blocos passou a formar outro supercontinente: o continente de Gondwana. Ele aglutinava as atuais América do Sul, África, Antártica, Austrália e Índia. Os blocos que formariam o Brasil começaram a se aproximar do desenho de hoje quando surgiu o último dos grandes continentes: a Pangea, há cerca de 300 milhões de anos. Foi da fragmentação de Pangea que nasceram os atuais oceanos e continentes, há aproximadamente 100 milhões de anos (Bicudo, 2002).
A história extraída das rochas reforça as hipóteses sobre a aparência que tinha a Terra há 1 bilhão de anos, um quinto de sua idade. Acredita-se que na época de Rodínia o planeta era uma imensa bola de gelo, resultante, provavelmente, de interferências astronômicas e alterações de órbita: o gelo aumentava a reflexão da luz, o que diminuía a absorção de energia. Veio, em seguida, o reverso da medalha: a intensa atividade vulcânica daquele período emitia uma quantidade enorme de gases, que acabaram por originar um gigantesco efeito estufa. O gelo começou a derreter e, em cerca de 10 mil anos, período geológico extremamente curto, a temperatura da Terra passou de 50ºC (graus Celsius) negativos para 50ºC positivos. E isso causou um impacto tremendo na vida existente naquela época especialmente na vida multicelular marinha (Bicudo, 2002).
A concha nacarada surgiu também neste Período, em resultado de múltiplos episódios de evolução convergente e como resposta ao aumento de predação resultante do GOBE (Vendrasco et al., 2011, 2013). Registe-se que estudos realizados sobre a microestrutura da concha de Fordilla e Pojetaia demonstraram que esses bivalves primitivos não possuíam nácar nas suas conchas, mas sim aragonita foliada (Vendrasco et al., 2011).

Paleogeografia durante o início do Ordoviciano (circa 480 Ma). Da divisão do supercontinente Rodínia resultaram os continentes Gondwana, China do Sul, Laurêntia, Báltica e Sibéria. Dos processos de rifting nas margens da Gondwana resultaram uma série de novos territórios e microcontinentes, como a Avalónia [Fonte: adaptado de Cocks & Torsvik, 2002].
O principal momento de radiação terá ocorrido no início do Ordovícico, envolvendo a evolução das brânquias com funções alimentares (Cope & Babin, 1999). Esta alteração morfológica e fisiológica terá promovido o aparecimento de estratégias alimentares de maior sucesso, permitindo aos bivalves o aproveitamento do crescente suplemento de nutrientes disponíveis nos habitats, resultante das condições paleoambientais e paleogegráficas peculiares do Ordovícico.
Este passo evolutivo permitiu um rápido crescimento e diversificação, levando à ocupação de muitos novos nichos ecológicos e habitats, com os bivalves a percorrerem todo um leque de estratégias de infaunal a epifaunal.

Estilos de vida infaunal, semi-infaunal e epifaunal interpretados para alguns fósseis bivalves do Ordoviciano tardio encontrados no estado norteamericano do Kentucky (modificado de Pojeta, 1971, Fig. 9, p. 33). Bivalve lifestyle and ecology

Espécies animais do bento

Predaores do bento

Perfil do fundo do mar mostrando organismos típicos da infauna (de Tardent).
a) Cracas (Balaniden), b) Mexilhões azuis (Mytilus edulis), c) Poliqueta (Lanice conchilega), d) Poliqueta (Lagis koreni), e) Caracol (Littorina littorea), f) Molusco (Ensis americanus), g) Bivalve (Cerastoderma edule), h ) Bivalve (Scrobicularia plana), i) Bivalve (Mya arenaria), k) Poliqueta (Arenicola marina), l) Poliqueta (Hediste diversicolor), m) Bivalve (Macoma balthica)(Senckenberg).
As faunas de moluscos bivalves ordovicianos, mais precoces, estariam confinadas às plataformas de natureza siliciclástica da Gondwana e Avalónia (Babin, 2000; Cope, 2002; Cope & Kříž, 2013). No entanto, pelo Ordoviciano Médio, os bivalves já apresentavam uma diversidade claramente marcada pela latitude, facto que os passou a caracterizar até à modernidade.
Os Pteriomorphia (uma infraclasse de bivalves) distribuíam-se preferencialmente nas baixas latitudes, enquanto os Palaeoheterodonta e Heterodonta manifestavam preferência por latitudes mais elevadas.
No Ordoviciano tardio, os bivalves colonizaram as plataformas carbonatadas da Laurêntia e de Báltica, ocorrendo um segundo episódio de maior diversificação, principalmente em relação aos bivalves pteriomorfos (concha da Shel oil, que inclui diversas ordens entre elas a ordem Peticnidae).
Antes do final do Ordoviciano, todas as subclasses já se encontravam estabelecidas e a classe Bivalvia dispersara-se pela generalidade dos ambientes marinhos ao tempo existentes (Babin, 2000).
O final do Período Ordoviciano é marcado pela extinção hirnantiana, o primeiro dos cinco megaeventos de extinção de primeira ordem do Fanerozoico e, também, o primeiro a envolver paleocomunidades estruturadas em metazoários (Sepkoski, 1996).
Este evento compreendeu duas fases de extinção, em que à primeira se associam mecanismos como o arrefecimento induzido por importantes episódios de glaciação e descida do nível médio dos oceanos (e.g. Sutcliffe et al., 2000), bem como por alterações significativas nos seus ciclos geoquímicos (e.g. Zhang et al., 2009).
Os eventos de extinção Ordoviciano-Siluriano, também conhecidos como extinção em massa do Ordoviciano Tardio (LOME: Late Ordovician Mass Extinction), são coletivamente o segundo maior dos cinco maiores eventos de extinção na história da Terra em termos de porcentagem de gêneros que se tornaram extintos.
A extinção foi global durante este período, eliminando 49-60% dos gêneros marinhos e quase 85% das espécies marinhas. Apenas a extinção em massa do Permiano-Triássico excede o LOME em perda total de biodiversidade.
O evento de extinção afetou abruptamente todos os principais grupos taxonômicos e causou o desaparecimento de um terço de todos os braquiópodes e briozoários. Famílias, bem como numerosos grupos de conodontes, trilobitas, equinodermos, corais, bivalves e graptólitos. Essa extinção foi o primeiro dos "cinco grandes" eventos de extinção em massa fanerozóico e foi o primeiro a afetar significativamente as comunidades baseadas em animais. No entanto, o LOME não produziu grandes mudanças nas estruturas do ecossistema em comparação com outras extinções em massa, nem levou a nenhuma inovação morfológica específica. A diversidade gradualmente recuperou-se para níveis pré-extinção ao longo dos primeiros 5 milhões de anos do período Siluriano.
A extinção em massa do Ordoviciano Tardio é geralmente considerada como ocorrendo em dois pulsos distintos.
O primeiro pulso começou na fronteira entre os estágios Katiano e Hirnantiano do Período Ordoviciano Superior.
Este pulso de extinção é tipicamente atribuído à glaciação do Ordoviciano Superior, que se expandiu abruptamente sobre o Gondwana no início do Hirnantiano e mudou o clima da Terra de uma estufa para uma casa de gelo.
O resfriamento e a queda do nível do mar provocados pela glaciação levaram à perda de habitat para muitos organismos ao longo das plataformas continentais, especialmente taxa endêmica com tolerância restrita de temperatura.
Durante esse pulso de extinção, também ocorreram várias mudanças marcantes nos isótopos de carbono e oxigênio biologicamente responsivos. A vida marinha foi parcialmente rediversificada durante o período frio e um novo ecossistema de água fria, a "biota de Hirnantia", estabeleceu-se.
Anóxia e euxínia
Outro fator amplamente discutido na extinção em massa do Ordoviciano final é a anóxia, a ausência de oxigênio dissolvido na água do mar. A anóxia não apenas priva a maioria das formas de vida de um componente vital da respiração, mas também estimula a formação de íons metálicos tóxicos e outros compostos.
Um dos mais comuns desses produtos químicos tóxicos é o sulfeto de hidrogênio, H2S) um resíduo biológico e principal componente do ciclo do enxofre. A depleção de oxigênio, quando combinada com altos níveis de sulfeto, é chamada de euxínia. Embora menos tóxico, o ferro ferroso (Fe 2+) é outra substância que se forma comumente em águas anóxicas.
A anóxia é o culpado mais comum para o segundo pulso do LOME e está conectada a muitas outras extinções em massa ao longo do tempo geológico. Ele também pode ter desempenhado um papel no primeiro pulso do LOME, embora o suporte para essa hipótese seja inconclusivo e contradiga outras evidências de altos níveis de oxigênio na água do mar durante a glaciação.
O segundo intervalo de extinção está relacionado com um evento anóxico oceânico ocorrido na sequência de uma transgressão eustática iniciada durante o Hirnantiano terminal (e.g. Vandenbroucke et al., 2015).
No entanto, esta grande extinção poderá ter resultado de um conjunto de fatores desencadeantes, incluindo uma euxínia generalizada em conjunto com a destruição de habitats, devido às reconfigurações resultantes dos movimentos das placas tectónicas (Harper et al., 2014).
Segundo Cope (2002), a diminuição do nível eustático à escala global terá exposto as plataformas carbonatadas de baixa latitude, resultando na extinção de muitos géneros de bivalves epi e semi-infaunais, incluindo os únicos bivalves perfuradores do Ordovícico. Após a extinção hirnantiana, o Silúrico trouxe uma importante reposição ao nível das categorias taxonómicas da família e do género, de tal forma que as faunas silúricas se tornaram cosmopolitas, encontrando-se em plataformas com águas oxigenadas de todas as latitudes (Babin, 2000). Esta dispersão teria sido devida ao restabelecimento generalizado do sistema de correntes oceânicas, o qual permitiu o aparecimento de habitats de fundo marinho favoráveis nas latitudes mais elevadas da peri-Gondwana e Sibéria, colonizáveis por larvas provenientes das regiões tropicais da Laurentia e Báltica (Kříž, 2001; Cope & Kříž, 2013). Durante o Período Silúrico e o início do Devónico, os Pteriomorphia silúricos da peri-Gondwana (e.g. Slavidae, Cardiolidae, Praecardiidae, Buchiolidae, Stolidotidae, Spanilidae), essencialmente adaptados a um modo de vida bissado nas fácies calcárias, experimentaram também uma rápida evolução à escala geológica (Kříž, 2007), dando origem a várias linhagens envolvendo adaptações e modos de vida infaunais, semi-infaunais e epifaunais. Estes bivalves extinguiram-se, provavelmente, no Carbonífero inicial e não desempenharam qualquer papel na futura evolução desta classe de moluscos (Kříž, 2007).
Durante o Devoniano, para além da diversificação em curso desde o Siluriano, regista-se a primeira colonização de habitats de água doce (Babin, 2000) que se tornaria ubíqua durante o Carbonífero, particularmente nos paleoambientes de natureza deltaica.
São herdeiros dessas linhagens, que colonizaram os ambientes dulceaquícolas, os atuais Unionoida (Palaeoheterodonta) e algumas famílias de heterodontes (i.e., Corbiculidae, Dreissenidae e Sphaeriidae), bastante comuns em cursos de água na europa pricipalmente em Portugal, com exceção da penúltima, apesar de existirem evidências da sua presença no paleo-Tejo plio-plistocénico.
A diversificação de bivalves durante o Devoniano levaria a que vários taxa atingissem o seu acme (estágio culminante, ponto mais alto; clímax, auge, culminância) entre o Devoniano Inferior e o Devoniano Médio (e.g. Actinodesma, Gosseletia, Pseuda-viculopecten, Paracyclas, Cardiola e Buchiola), tendo entrado em forte declínio durante a transição Frasniano-Famenniano e tornando-se virtualmente extintos no final do Devoniano (Amler, 1999).
No entanto, segundo Amler (1999), uma nova e importante diversificação de bivalves ter-se-á ainda iniciado durante a transgressão eustática que caracterizou o final do Famenniano, dando origem a vários novos taxa que exibem já caracteres modernos e atravessam o limite Devoniano-Carbonífero, gerando as linhagens que ainda hoje conhecemos.
Durante o Triássico, logo após a extinção do final do Permiano, assistiu-se a um incremento do número de géneros de bivalves, com um auge no Carniano e predominío das espécies com características infaunais (McRoberts, 2001).
Este recurso à proteção propiciada por substratos móveis, surge, provavelmente, como resposta antipredação ao evento MMR (Mesozoic Marine Revolution) (Vermeij, 1977). Mais tarde, ainda durante o Mesozoico, algumas linhagens principais extinguiram-se, como foi o caso dos rudistas. Estes bivalves epifaunais de vida séssil colonizaram as águas pouco profundas desde o Jurássico tardio até ao final do Cretácico, onde foram corresponsáveis pelo desenvolvimento de edifícios recifais (Skelton & Smith, 2000).
A revolução marinha mesozóica foi o grande aumento dos predadores do fundo do mar que comiam moluscos. O termo foi usado por Vermeij, um paleontólogo que passou anos investigando mudanças nos invertebrados do fundo do mar.
Houve uma mudança notável na vida no fundo do mar entre a fauna paleozóica e a fauna moderna. Esta mudança ocorreu durante o Mesozóico. Na era Mesozóica, os predadores do fundo do mar desenvolveram várias novas formas que se alimentavam dos abundantes mariscos (brachiopods e bivalves). Estes predadores ainda são abundantes hoje em dia: são principalmente as estrelas-do-mar, os gastrópodes e os caranguejos.
Cada predador tem seus próprios métodos. Os caranguejos quebram as conchas pela força. Vários tipos de gastrópodes desenvolveram a capacidade de entrar em conchas. Os Muricidae perfuram através das conchas e consomem presas. Alguns gastrópodes fazem buracos na concha e colocam uma substância paralisante ou relaxante; outros trabalham em pequenas rachaduras na borda da concha. Uma vez que entram com probóscide, eles comem os moluscos. Talvez surpreendentemente, algumas esponjas também possam perfurar as conchas (Alegsa, 2021).
Classe Aplacophora
São moluscos pouco conhecidos, vermiformes que não apresentam concha. Podem ter de 2,5 a 20,0 cm de comprimento, e podem ser epibentôncos e infaunais. Algumas espécies são capazes de viver em grandes profundidades.
Apresentam espículas ou escamas calcárias distribuídas pelo tegumento. Estas escamas ou espículas são produzidas pelas glândulas da concha dispersas na epiderme.
Os moluscos são o grupo irmão de Annelida, por isso esses grupos basais apresentam um formato vermiforme e uma larva trocófora. Por isso estão agrupados no clado Lofotrocozoa.


Aplacóforo Neomeniomorpha Amboherpia heterotecta.

Solennogaster Epimenia spp. Apresenta uma cutícula calcária e aragonítica.
Apresentam uma boca anterior com cirros e papilas sensoriais
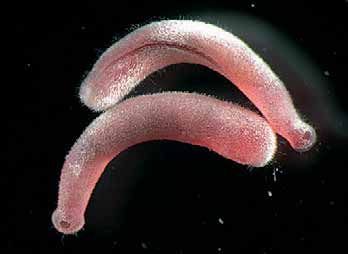
Aplacóforos
CLASSE POLYPLACOPHORA
O nome dessa classe vem do fato de possuírem muitas placas muitas placas em sua concha. Polyplacophora, do grego poly: muitos + placa: placas + phere: possuir, portar.
Possuem sete placas nas formas jovens e oito nas formas adultas.
Os poliplacóforos são popularmente chamados de quítons e vivem aderidos a rochas nas regiões entre marés ou submersas em costões rochosos e até em grandes profundidades. Sua concha é formada por uma série de oito placas sobrepostas e articuladas. Essas placas são unidas por um cinturão (parte do manto recoberta por espessa cutícula que contorna as placas.
Os Polyplacophora têm corpo de formato oval, alongado e achatado dorso-ventralmente, com uma concha composta de oito placas dorsais, independentes, mas articuladas. Entre as placas imbricadas apresentam células epiteliais que atravessam a concha chamadas de estetos. Os estetos são estruturas específicas dos quítons, que podem ser tanto sensoriais quanto secretoras. Os estetos consistem em um agrupamento de células especializadas e podem ser classificados como microestetos e macroestetos. Estão localizados em canais verticais nos tegumentos das valvas, forrados por epiderme. Alguns estetos apresentam ocelos, associados à fotorrecepção.
O pé forma uma sola rastejadora achatada e ventral. A sola do pé é ciliada e repleta de células glandulares que produzem muco para o deslocamento e fixação e assim deixam uma trilha de muco por onde o animal passou.
Quítons também utilizam seu amplo cinturão para aderir ao substrato, firmando-se fortemente e levantando a margem interna do cinturão, de forma a criar um ligeiro vácuo.

Algumas características morfológicas de um quíton

Quíton


Terminologia para válvulas de quíton. Observe que há alguma sobreposição e gradação na terminologia da escultura tegmental. Por exemplo, "treliça" por definição contém "estrias longitudinais". Além disso, as saliências pequenas e estreitamente espaçadas rotuladas como "grânulos" se transformam em saliências maiores e mais espaçadas rotuladas como "tubérculos". Veja Schwabe (2010) para mais detalhes sobre a terminologia do Quíton (Vendrasco e cols, 2012).

by G. J. Browning from Marine Molluscs of Victoria (GJB)

CLASSE MONOPLACOPHORA
Os monoplacóforos são exclusivamente marinhos e habitam águas profundas. Seu corpo é totalmente recoberto por uma concha única, vindo daí seu nome, do grego mono: um.
Monoplacóforos (Monoplacophora) são uma classe de moluscos ancestrais que foram considerados extintos, já que o grupo era conhecido apenas através de fósseis do período cambriano e devoniano, até que em 1952 foram encontrados espécimes vivos do gênero Neopilina, extraídos de uma cova subaquática profunda nas costas da Costa Rica na América Central.
Desde que Neopilina foi descoberta em 1952, espécimes de Monoplacophora de diferentes espécies foram coletados, mas sempre em águas profundas (2.000 a 7.000 m de profundidade).
As espécies de Neopilina foram coletadas no Oceano Pacífico Oriental, no Oceano Atlântico Sul e no Golfo de Áden. O grupo Monoplacophora é considerado o grupo basal do qual derivam os moluscos das classes Gastropoda, Bivalvia e Cephalopoda.
Os monoplacóforos são moluscos de simetria bilateral com uma "pseudometameria", não se pode dizer que apresentam metameria, mas seus órgãos apesentam-se como múltiplos pares, por exemplo: múltiplos pares de músculos retratores do pé, múltiplos pares de nefrídios, múltiplos pares de ctenídios (brânquias)etc.
Possuem uma concha simples do tipo pateliforme ou escudo que cobre o corpo e se estende completamente sobre a parte de trás do animal. Em geral, Monoplacophora se assemelha a Polyplacophora, mas com seis pares de nefrídios dispostos metamericamente, que descarregam detritos por nefridióporos associados a cinco ou seis pares de brânquias simples (dependendo da espécie), dispostos alternadamente com oito pares músculos dorsoventrais, que estão ligados à concha e ao pé.
O celoma, consiste de uma cavidade pericárdica caudal e na qual existem dois pares de grandes gônadas ventrais.
O sistema circulatório consiste principalmente em átrios pares que drenam para os ventrículos (também pares), um de cada lado do intestino.
O sistema digestório é semelhante ao observado no restante dos moluscos.
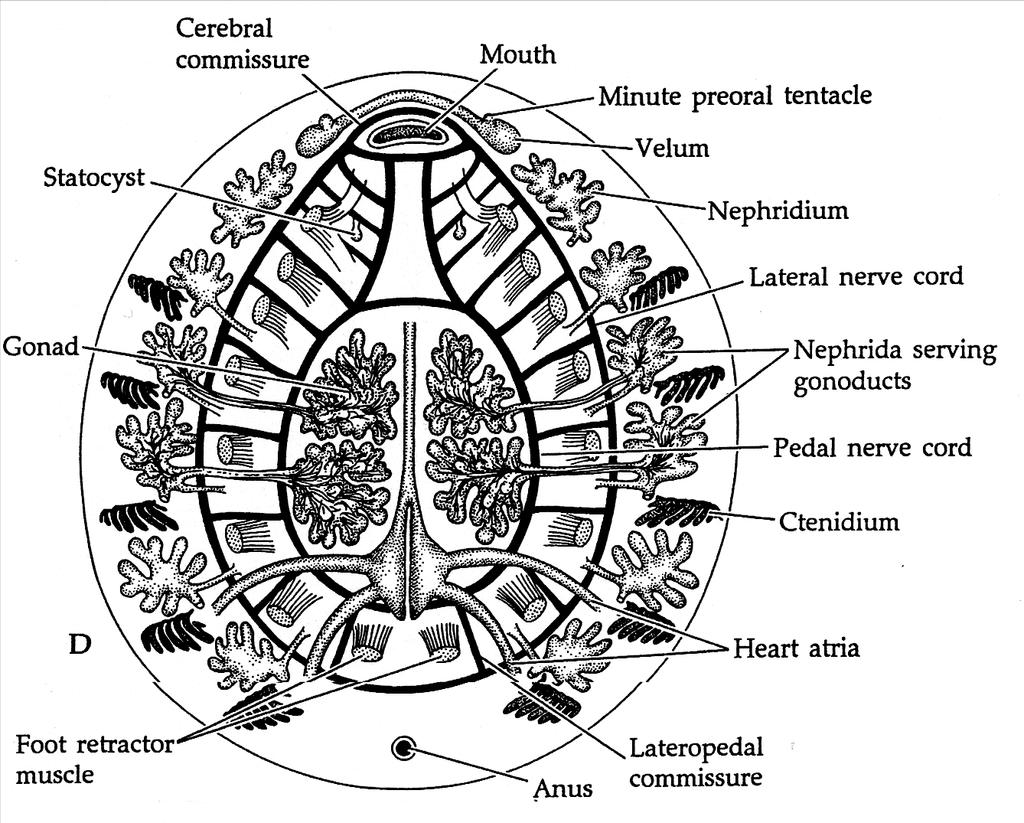
Anatomia de um monoplacófora
O sistema reprodutivo consiste em dois pares de gônadas grandes (em cada gonocele, cavidade) que fluem para o terceiro e quarto nefridióporos, respectivamente, com os gametas saindo pelas aberturas do nefrídio.



Monoplacófora, Pilina ungis

Monoplacófora, Triblidium reticulatum

Tryblidium reticulatum, a typical Early Palaeozoic shallow-water Tryblidiid.
Length about 3.5 cm. Silurian (Wenlock Age) of Baltica (Gotland);
Monoplacophora - Tryblidiida - Tryblidiidae
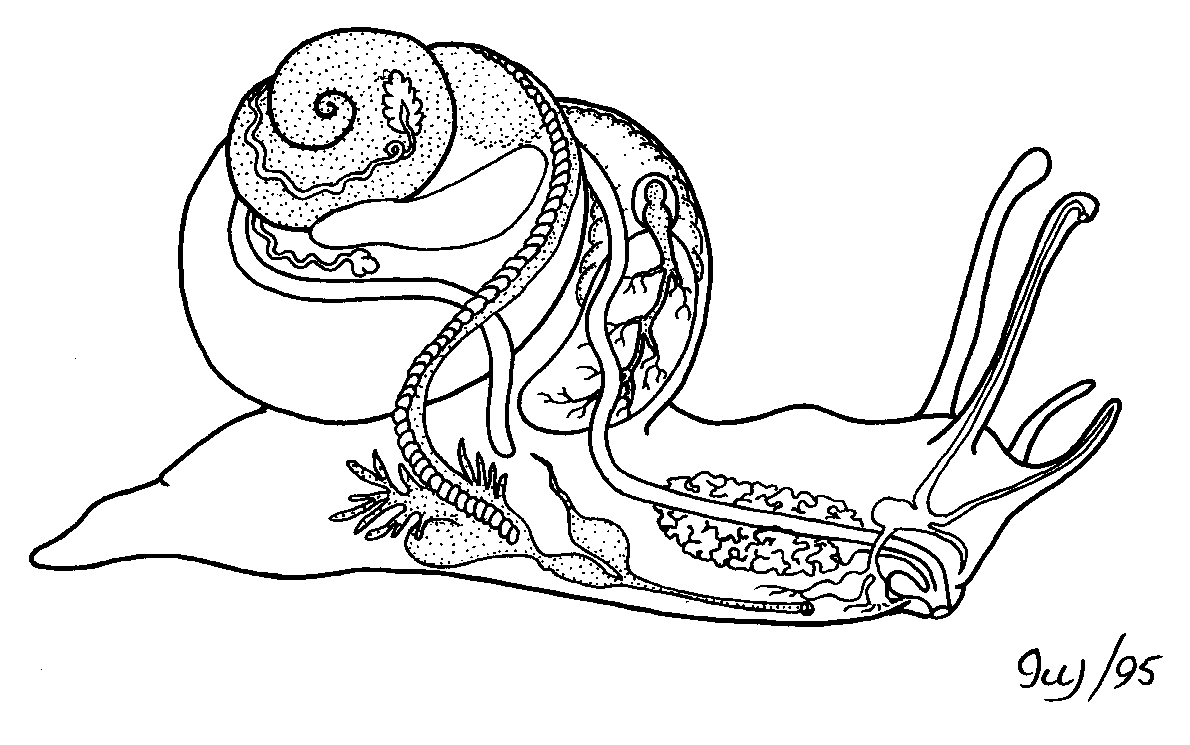
CLASSE GASTROPODA
É a que apresenta o maior número de espécies e maior diversidade de habitats entre os moluscos, sendo a única com representantes em três tipos de ambiente: marinho, de água doce e de terra firme.
Alguns gastrópodes bem conhecidos são as lesmas, os caracóis de jardim e os caramujos.
Existem espécies com concha, geralmente espiralada, como os caracóis e caramujos, ou sem, como as lesmas.
O nome do grupo faz referência à localização de seu pé ventral, do grego antigo γαστήρ (gaster) + πούς (pous): estômago+pés = gastrópode.
O pé é ventral e musculoso, situado abaixo do estômago, uma verdadeira sola rastejadora.
Os gastrópodes são o grupo irmão dos cefalópodes. A linha evolutiva foi mais ou menos desta forma:
Uma característica importantíssima dos gastrópodes é a torção. E evidências fósseis e do desenvolvimento mostram que a torção ocorreu da seguinte forma:
Cavidade do manto lateral - Monoplacófora, poliplacófora e monoplacófora
Cavidade do manto posterior - arquétipo
cavidade do manto anterior - gastrópoda
Esta característica é chamada de torção. A torção trouxe a cavidade do manto da região posterior para a região anterior, o que gerou um grau de auto-poluição para o organismo. Ou seja, a abertura do sistema digestório e excretor ficou bem acima da cabeça, o que ocasiona a eliminação de resíduos sobre as branquias e sobre estruturas sensoriais. Para minimizar esse problema as estruturas do lado esquerdo desapareceram.
Sistema nervoso
ANEL NERVOSO AO REDOR DO ESÔFAGO
COM DOIS CORDÕES NERVOSOS: UM VISCERAL E OUTRO PEDIOSO
Moluscos gastrópodes devido a torção apresentam uma inversão da cavidade do manto em relação ao eixo cabaça-pé; com isso, a porção posterior do sistema nervoso está dobrada e torcida em forma de 8 condição conhecida como estreptoneura.
A torção trouxe os gânglios nervosos posteriores para região anterior do animal.
Moluscos destorcidos – sistema nervoso exibe uma simetria bilateral derivada, com cordões nervosos viscerais mais ou menos retos e paralelos – condição conhecida de eutineura.
Órgãos Sensoriais
Maioria dos gastrópodes apresentam 1 ou 2 pares de tentáculos cefálicos;
Os gastrópodes pulmonados – possuem tentáculos ópticos e apresentam olhos nas extremidades destes.
Arqueogastropodes – tentáculos epipodiais curtos nas bordas do pé ou do manto;
Tentáculos cefálicos podem ser dotados de olhos, células táteis e quimiorreceptores.
Opistobrânquios – tem na região dorsal anterior um par de tentáculos quimiorreceptores ramificados ou pregueados chamados de rinóforos. Opistobrânquios – apresentam dois pares de tentáculos cefálicos
Reprodução
Gastrópodes podem ser dioicos e monoicos
Desenvolvimento pode ser direto, indireto ou misto
Desenvolvimento indireto – larva trocófora livre-nadante.
Em alguns grupos de moluscos a trocófora é seguida de um estágio larval exclusivo dos moluscos – larva véliger.
Larva véliger possui um pé, concha, opérculo e outras estruturas semelhantes às dos adultos.
Larva trocófora - larva véliger – juvenil - adulto



Conchas de Fluminicola modoci
Shells of UKL F. modoci (A-G) and F. fresti (H) A USNM 1144454 B USNM 1144520 C USNM 1144942 D USNM 1144407 E USNM 1144966 F USNM 1190096 G USNM 1144565 H USNM 1144900. Scale bar: 1.0 mm.
Amnicola dali
(Fonte WikiMili)

Caramujo, um gastrópode terrestre


Gastrópode terrestre (lesma verdadeira) note a abertura do poro respiratório

Amarelo claro - partes moles
Castanho (verde oliva) - concha e opérculo
Verde - sistema digestório
Lilás - brânquias
Amarelo - osfrádio (órgão quimossensorial)
Vermelho - coração (sistema hemal)
Rosa - cavidade pericárdica
Violeta - Gônada
1. Pé (parte anterior da sola rastejadora)
2. Gânglio cerebral
3. Pneumóstoma (poro respiratório ou abertura do sistema respiratório)
4. Alta comissura (cordão nervoso visceral)
5. Osfrádio (órgão quimossensorial)
6. Brânquias
7. Gânglio pleural
8. Átrio do coração
9. Gânglio visceral
10. Ventrículo
11. Pé
12. "Tampa" do opérculo
13. Cérebro
14. Boca
15. Tentáculo (quimiossensorial, 2 ou 4)
16. Olho
17. Pênis (evertido, normalmente interno)
18. Anel nervoso esofágico
19. Gânglio pedal (gânglio que lida com a inervação do pé)
20. Baixa comissura
21. Vaso deferente (canal que leva espermatozoide para o pênis)
22. Cavidade palial / cavidade do manto / cavidade respiratória (nos pulmonados)
23. Gânglio parietal
24. Ânus
25. Hepatopâncreas
26. Gônada
27. Reto
28. Nefrídio

2. Fígado
3. Pulmão
4. Ânus
5. Poro respiratório (pneumóstoma)
6. Olho
7. Tentáculo
8. Gânglios cerebrais
9. Ducto salival
10. Boca
11. Esôfago
12. Glândula salival
13. Poro genital
14. Pênis
15. Vagina
16. Glândula mucosa
17. Oviducto
18. Saco de dardos
19. Pé
20. Estômago
21. Rim (metanefrídio)
22. Manto
23. Coração
24. Vasos deferentes (oriundos da gônada).
Aqueles que perderam as brânquias as trocas gasosas são realizadas através do manto.
Gastrópodes exclusivamente terrestres não possuem ctenídios.
Trocas gasosas ocorrem através de regiões vascularizadas do manto, localizadas na cavidade paleal.
A designação de pulmão está relacionado a organização desta cavidade.
Nos pulmonados terrestres, as margens do manto estão fundidas ao dorso do animal, selando a cavidade do manto, exceto uma pequena abertura do lado direito chamada de pneumóstoma.









](http://www.caledonianseashells.com/img/p/4239-17264-thickbox.jpg)




SISTEMA CIRCULATÓRIO
Os gastrópodes apresentam um sistema circulatório do tipo aberto que utiliza como fluido de transporte para o s gases a hemolinfa. Na vasta maioria das espécies na hemolinfa está presente o pigmento respiratório hemocianina, de cor azul, mas a família pulmonada Planorbidae tem a hemoglobina, pigmento de cor vermelha, como proteína respiratória.
Bebe caramujo saindo do ovo (eclosão de um gastrópode terrestre)





Megalobulimus spp gastrpodes terrestres nativos do Brasil
Gastrópode terrestre
SISTEMA EXCRETOR
O órgão excretor primário dos gastrópodes são os nefrídios, os quais produzem amônia ou ácido úrico como principal excreta. O funcionamento dos nefrídios também exerce um importante papel na manutenção do balanço hídrico corporal nas espécies de água doce e terrestres. Os moluscos terrestres excretam ureia e os moluscos aquáticos excretam amônia.
Cladograma mostrando as relações entre os gastrópoda,
para mostrar os táxons desta classe.
CLASSE CEPHALOPODA
Pés na cabeça
Moluscos sem concha externa, apresentando uma conha interna com câmaras.
A concha pode ser externa como no Nautilus, interna nas espírulas, sépias e lulas. A concha da sépia chama-se de pena ou gládio e apresenta câmaras, com as quais o animal regula sua densidade, podendo afundar ou subir na coluna d'água.
Estão nesta classe o polvo, a lula, o náutilo, as sépias e o calamar, animais exclusivamente marinhos.
O pé dos cefalópodes é dividido em tentáculos. Seu habitat é exclusivamente marinho.
Apresentam circulação é fechada com um coração sistêmico e um coração na base das brânquias.
Olhos (com cristalino, córnea) muito semelhantes ao dos mamíferos mas análogos a estes. Nestes animais o cristalino é movido para frente ou para trás, não é maleável como nos olhos dos vertebrados mamíferos.
Muitas espécies possuem células especiais e pigmentadas chamadas cromatóforos, que permitem a esses animais mudar de coloração. Alguns cefalópodes, como as lulas, também são dotados de um pequeno órgão, em forma de saco, que contém um fluído escuro e liberam pelo ânus. Quando ameaçados, esses animais liberam o fluído, dificultando a visão do predador.

CLASSE BIVALVIA OU PELECIPODA
Os bivalves são um grupo grande e muito diverso. è o segundo maior grupo de molusco. Existem espécies marinhas e de água doce. Alguns exemplos são as ostras, os mexilhões e os mariscos. Possuem uma concha composta por duas partes, chamadas de valvas, articuladas por uma espécie de dobradiça situada na região dorsal. Possuem uma cabeça reduzida e um pé estreito que lembra a forma de um machado.
Abertura da concha ligamento elástico (para abertura) e musculatura adutora (que puxa uma valva contra a outra).
São divididos em três grupos:
Protobrânquios
Vivem enterrados no sedimento
São comedores seletivos de depósitos
branquias simples e palpos ou tentáculos que revolvem o sedimento que sera usado como alimento.
Lamelibrânquia
São animais filtradores de suspensão
Brânquias muito alongadas relacionado com a alimentação
Tornaram independentes do substrato para a alimentação, não necessitando ficar enterrados; tornaram-se epibentônicos.
Algumas são bissadas (apresentam bisso) ou sedimentados por uma das conchas como a ostra ao substrato.
Pectem é livre e pode nadar curtas distâncias
Espécies perfuradoras de madeira
Septibrânquia
Bivalbes predadores
Sugam a presa para dentro da concha
Nodipecten fragosus (Conrad,1849).
Nodipecten fragosus (Conrad,1849).
Donax variabilis, de Folly Beach, Carolina do Sul (SC).
CLASSE SCAPHOPODA
As duas valvas da concha se fundem fromando uma estrutura semelhante a um dente
comedores seletivos de depósito
pé escavador, vivem enterrados com a extremidade posterior para fora
São moluscos exclusivamente marinhos cuja concha lembra uma pequena presa de elefante, oca e aberta nas duas extremidades. São popularmente conhecidos como concha-dente. O pé desses animais é musculoso e adaptado a escavar o substrato. Vivem parcialmente enterrados na areia. Conhecidos, em geral, por dentálios.
Antalis entalis (Scaphopoda: Dentaliida: Dentaliidae);
conchas vazias lavadas após tempestades;
Swanage Beach, Inglaterra.
Os escafópodes são chamados de conchas de dente ou presa porque alguns tipos se assemelham a presas em miniatura de elefante. Esses moluscos pouco conhecidos são na verdade sobreviventes de um grupo paleozóico mais diverso conhecido como Rostroconchia. Eles tem sido agrupados de forma diversa com bivalves e cefalópodes (inclusive sendo considerados como grupo irmão de bivalvia). Embora uma evidencia seja a estrutura da concha que é igual a dos demais chonchíferos.
RESUMO
Filo Mollusca
Representantes
Caracóis, lesmas, mexilhões, ostras, polvos, lulas, nautilus.
Segundo maior grupo em número de espécies (90 mil espécies vivas e 70 mil fósseis).
Habitat
Principalmente marinhos:
Epibentômicos, i.e., vivendo sobre o substrato, infaunais, i.e., enterrados no substrato e também podem perfurar madeira ou calcário; podem ser viver em água rasas ou em grandes profundidades, pelágicos, nadam nas profundezas.
A classe gastrópoda e bivalves invadiram o ambientes de água doce e os gastrópodes são os únicos que invadiram o ambiente terrestre.
Importância
Alimentação humana: polvo, lula, mexilhão, escargot
Joalheria: pérola e madrepérola
Agricultura: pragas agrícolas (caracóis herbívoros),
Medicina: vetores de doenças (esquistossomose) gastrópode Planorbideo.
A classe gastrópoda e bivalves invadiram o ambientes de água doce e os gastrópodes são os únicos que invadiram o ambiente terrestre.
Importância
Alimentação humana: polvo, lula, mexilhão, escargot
Joalheria: pérola e madrepérola
Agricultura: pragas agrícolas (caracóis herbívoros),
Medicina: vetores de doenças (esquistossomose) gastrópode Planorbideo.
Ecológica: reciclagem de nutrientes, polinização (malacofilia) gastropodes.
Plano corpóreo básico:
Simetria bilateral pois somente podemos passar um plano de corte através do corpo desses organismos.
Molusco arquétipo:
Plano corpóreo básico:
Simetria bilateral pois somente podemos passar um plano de corte através do corpo desses organismos.
Molusco arquétipo:
Corpo constituído por uma cabeça, por um pé e massa visceral. Algumas vezes a cabeça e o pé são referidos como massa céfalo-pediosa. Abaixo vemos um molusco arquetípico; esse esquema é importante para localizarmos os órgãos dentro do plano corporal desse filo. Como sabemos o arquétipo dos moluscos seria mais semelhante a um "verme" muito parecido com os moluscos da classe Aplacófora.

Molusco arquétipo
Quanto ao destino do blastóporo:
São organismos protostômios, i.e., o blastóporo dá origem a boca.
Cavidade do corpo:
Primeiros animais a apresentar celoma verdadeiro, embora este seja muito reduzido. O celoma dos moluscos restringe-se a regiões do coração, gônadas e nefrídios e parte do intestino. O celoma é do tipo esquizocélico, i.e., formado a partir de fendas internas que surgem da mesoderme do embrião, com preenchimento completo da blastocele por mesoderme seguido por um processo de morte celular, dando origem à cavidade celômica revestida completamente por mesoderme.
Sistema circulatório:
Sistema circulatório aberto ou lacunar, coração com 2 câmaras (um átrio e um ventrículo).
Manto:
O corpo é envolvido pelo Manto, espessa camada epidérmica-cuticular trilobada que produz 1 ou 2 valvas da concha calcária.
Clado monofilético:
Acredita-se que os moluscos constituam um grupo de organismos todos derivado de um ancestral comum.
Exercícios
1. Leia a reportagem sobre malacocultura no link (https://agro20.com.br/ostra/) e descreva a importância da ostra para economia e para o homem.
2. Cite a importância dos moluscos em geral para o ser humano.
3. Esquematize o arquétipo de um molusco e coloque o nome de suas partes.
4. Cite as características e inovações do filo Mollusca.
5. Complete a tabela (abaixo) e poste no seu blog
5. Complete a tabela (abaixo) e poste no seu blog


Rodínia
Acredita-se que este supercontinente formou-se há 1 bilhão de anos e que abrangia a maior parte da porção continental da Terra. Acredita-se que quebrou-se em oito continentes cerca de 750 milhões de anos atrás. Durante a época que existiu este super continente a Terra ficou toda congelada (hipótese da Terra bola de neve), com temperaturas muito baixas e com seu oceano tendo uma capa de gelo que poderia ter em torno de um quilômetro de profundidade. Antes do congelamento do super continente formou-se grande deserto sem vida vegetal ou animal. Os movimentos dos continentes antes da formação de Rodínia são incertos.
Entretanto os movimentos das massas continentais depois do rompimento do supercontinente são melhor compreendidos e continuam sendo objetos de pesquisa. Os oito continentes que compunham Rodínia foram posteriormente reunidos em outro supercontinente chamado Pannótia e, depois, Pangeia. Os vestígios de Rodínia podem ser encontrados pela América do Sul. No Brasil podem ser representados pelo cráton São Luís, pela província Borborema, pelo bloco Parnaíba, pelos crátons São Francisco e Paranapanema, pelo bloco Rio Apa, pelo cráton Luiz Alves, pelo maciço Curitiba, parte do cráton Rio da Plata, sendo o principal vestígio o cráton Amazônico que compreende vários estados do Norte do Brasil e parte da região Centro-Oeste.

Reconstrução proposta de Rodínia para 750 Ma, com cinturões orogênicos de idade de 1,1 Ga destacados em verde. Pontos vermelhos indicam granitos tipo A de 1,3–1,5 Ga.(Ga: bilhões de anos)(WP, Bicudo, 2002).

Rodinia a 900 Ma. "Reconstrução de Consenso" de Li et al. 2008.
Fonte
https://cesad.ufs.br/ORBI/public/uploadCatalago/11265816022012Biologia_dos_Invertebrados_I_aula_8.pdf
Bibliografia
STORER, T. e USINGER, R. Zoologia Geral. Rio de Janeiro, Campanha Ed. Nacional, 1978.
RUPPERT, E. E., BARNES R. D. Invertebrate Zoology. NYC, Saunders College Publ. (1995).
LOPES, S. e ROSSO, S. Biologia vol. 3 Ed. São Paulo, Ed. Saraiva. (2010)
MONGE-NÁJERA, J. Introducción: unvistazo a la historia natural de los moluscos. Rev. Biol. Trop., Suppl. 3, p. 1-3, 2003.
MORAES, Marcia Soman. Jornal da USP. O manual dos moluscos do Brasil. 19 nov. 2006
Ricardo J. Pimentel, Pedro M. Callapez & Paulo Legoinha. Um Éon de história dos Bivalvia: ideias sobre a sua origem, filogenia e importância paleontológica e educativa. in Abrantes, I., Callapez, P. M., Correia, G. P., Gomes, E., Lopes, B., Lopes, F. C., Pires, E., & Rola, A. (Eds.), Uma visão holística da Terra e do Espaço nas suas vertentes naturais e humanas. Homenagem à Professora Celeste Romualdo Gomes. Coimbra: CITEUC. CITEUC, 2020
(ótima fonte)
Tudo sobre Malacologia, moluscos, conchas etc...
Publicado em 12/VII/2020
Atualizado em 12/VIII/2021
Atualizado em 20/IV/2022
Atualizado em 03/I/2023
0 comentários:
Postar um comentário